Mikrobiyal Genetik
Bakteriler, ihtiyaç duydukları genetik değişkenliği nasıl elde ederler? Alışılmadık bir besin kaynağını parçalamak veya onları yok etmekle tehdit eden bir antibiyotiği bozmak için yeni bir gene ihtiyaç duyabilirler - bu geni edinmek yaşamla ölüm arasındaki fark anlamına gelebilir. Peki ama bu genler nereden geliyor? Bakteriler onları nasıl ele geçirmiş olabilir? Bakterilerin yeni genler edinmek için kullandıkları süreçleri yani Yatay Gen Transferi (HGT) olarak bilinen mekanizmaları keşfedeceğiz.
Konjugasyon
Konjugasyon, bir donör bakterinin bir plazmidin kopyasını bir pilus aracılığıyla bir alıcı bakteriye aktarması sürecidir. Bu süreç hücreden hücreye temas gerektirir. Donör hücre (F+), pilus olarak bilinen ipliksi bir filament yapmak için gerekli proteinleri kodlayan ekstrakromozomal bir dsDNA parçası olan konjugatif bir plazmide sahiptir. Pilus, alıcı (F-) hücreye bağlanarak onu donör hücreye yakınlaştırmak için kullanılır. Daha sonra iki hücre arasında bir kanal açılarak plazmidin bir ssDNA kopyasının alıcı hücrelere girmesine izin verildiğine inanılmaktadır. Her iki hücre de ardışık tek zincirli DNA'nın tamamlayıcı kopyasını yapar ve sonuçta konjugasyona uygun iki F+ hücresi oluşur.

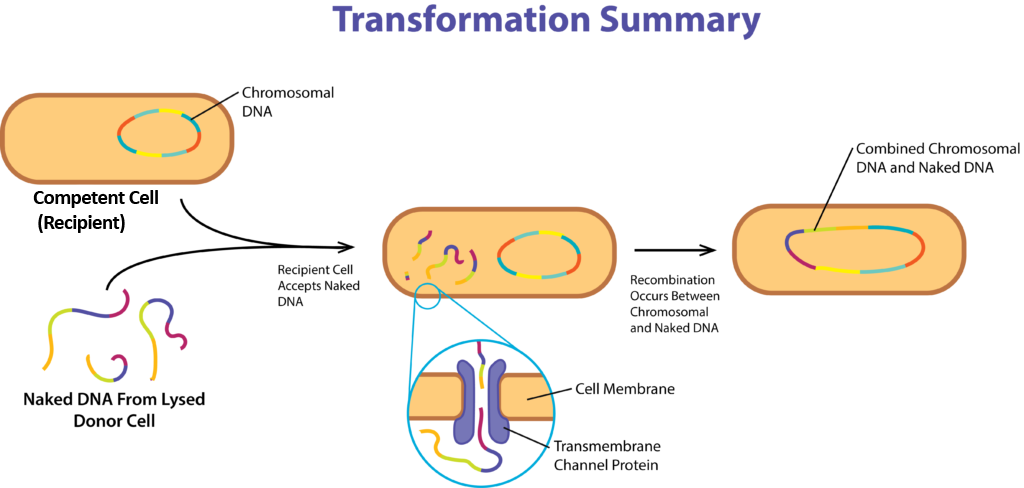
Transformasyon
Transformasyon süreci de bir bakteri hücresinin yeni genler edinmesini sağlar, ancak hücreden hücreye temas gerektirmez. Bu süreçte yeni genler doğrudan çevreden edinilir. Tipik olarak süreç, bir noktada parçalanmış ve çevreye çıplak DNA salmış bir donör hücre gerektirir. Alıcı hücre, DNA'yı çevreden alıp kendi genomuna dahil edebilen bir hücredir ve burada hücre kompetan olarak tanımlanır.
Bir hücreyi çevreden DNA almaya teşvik etmenin mekanik ve kimyasal yolları vardır, ancak doğal yeterlilik genetik olarak belirlenir. Bu süreç tipik olarak büyümenin üstel fazının sonunda veya durağan fazın başında, yüksek hücre yoğunluğu ve sınırlı besin varlığında gerçekleşir. Bu koşullar altında DNA bağlayıcı proteinler (DNA translokaz), endonükleazlar ve transmembran kanal proteinleri dahil olmak üzere spesifik proteinler üretilir. Gram negatif hücreler de DNA'yı dış zar boyunca taşımak için bir hücre duvarı otolizini yaparlar.
Rastgele DNA parçaları hücrenin dışındaki reseptörlere bağlanır ve daha sonra DNA translokaz tarafından, genellikle çok sayıda farklı proteini içeren büyük bir yapı olan transmembran kanalı yoluyla hücre içine taşınır. Hücreye sadece ssDNA geçebiliyorsa, dsDNA'nın bir ipliğini bozmak veya DNA parçasını daha küçük boyutlara bölmek için bir endonükleaz kullanılabilir. Hücre içine girdikten sonra, genlerin ifade edilebilmesi için DNA'nın RecA tarafından bakteriyel kromozoma dahil edilmesi gerekir (aşağıdaki Moleküler Rekombinasyon bölümüne bakınız).
Kalıt Aktarım (Transdüksiyon)
Transdüksiyon, bakteri genlerini bir hücreden diğerine aktarmak için bir kanal görevi görmek üzere bir virüsün, bir bakteriyofajın kullanılmasını içerir ve böylece hücreden hücreye temas gerekliliğini ortadan kaldırır. İki farklı transdüksiyon türü vardır: genelleştirilmiş transdüksiyon ve özelleşmiş transdüksiyon.
Genelleştirilmiş Transdüksiyon
Genelleştirilmiş transdüksiyonda, bir bakteriyel konak hücre, litik replikasyon döngüsüne giren virülan ya da temperat bir bakteriyofaj ile enfekte olur. Replikasyonun ilk üç aşamasından sonra (absorpsiyon, penetrasyon ve sentez), virüs, tam olarak oluşturulmuş viryonların yapıldığı birleşme aşamasına girer. Bu aşamada, bakteriyel DNA'nın rastgele parçaları yanlışlıkla bir faj kafasına paketlenir ve bu da bir transdüksiyon partikülünün üretilmesiyle sonuçlanır. Bu partiküller geleneksel anlamda bir hücreyi enfekte edemeseler de, yeni bir bakteriyel konakçı hücreye bağlanabilir ve DNA'larını içine enjekte edebilirler. DNA (ilk bakteriyel konak hücreden) alıcının kromozomuna dahil edilirse, genler ifade edilebilir.

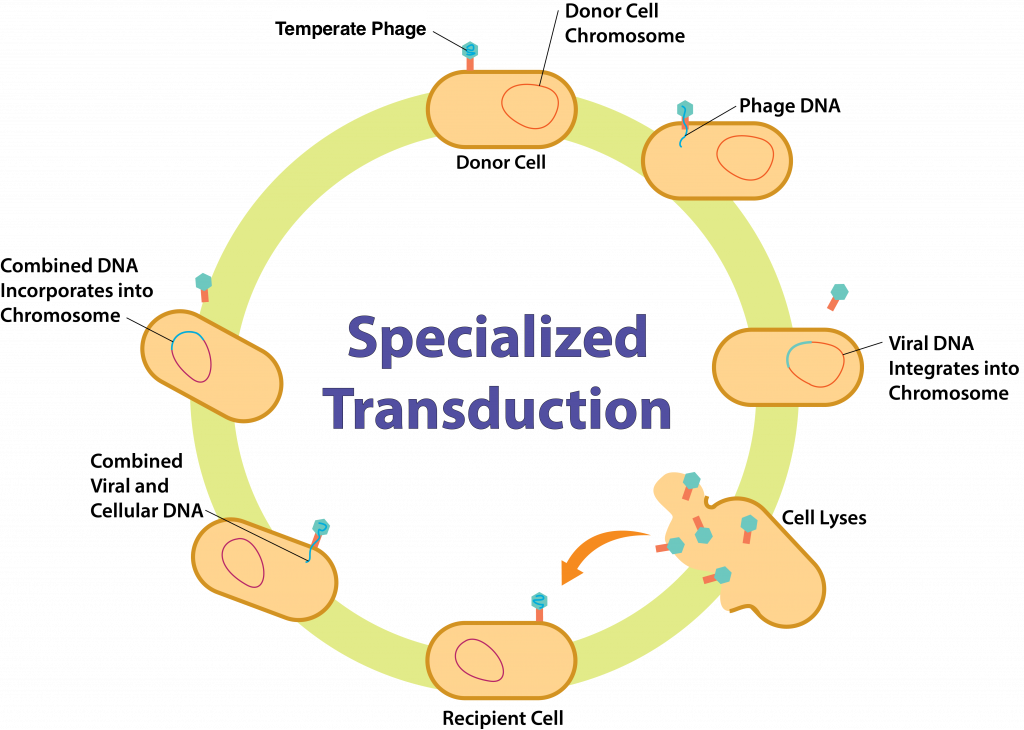
Özelleşmiş Transdüksiyon
Özelleşmiş transdüksiyon, lizojenik replikasyon döngüsünü içerdiğinden yalnızca temperat bakteriyofaj ile gerçekleşebilir. Bakteriyofaj rastgele bir bakteriyel konakçı hücreye bağlanarak viral DNA'yı içine enjekte eder. DNA, konak hücrenin kromozomuna entegre olarak bir profaj oluşturur. Bir noktada, profajın bakteriyel krozomdan çıkarıldığı indüksiyon meydana gelir. Özelleşmiş transdüksiyonda, eksizyon yanlış yapılır ve viral genlerin hemen bitişiğindeki bakteriyel genlerin bir kısmı da eksize edilir. Bu DNA sentez aşaması için şablon olarak kullanıldığından, tüm kopyalar viral ve bakteriyel DNA'nın bir melezi olacak ve ortaya çıkan tüm virionlar hem viral hem de bakteriyel DNA içerecektir.
Hücre parçalandıktan sonra virionlar diğer bakteriyel konak hücreleri enfekte etmek üzere serbest kalır. Her virion, konak hücreye bağlanacak ve DNA hibridini içeri enjekte edecektir; eğer bir proflaj oluşursa, bu, konak kromozomuna dahil edilebilir. Bu noktada ikinci bakteriyel konak hücre kendi DNA'sını, önceki bakteriyel konak hücreden gelen DNA'yı ve viral DNA'yı içerebilir.
Moleküler Rekombinasyon
HGT vakalarının her birinde, süreç ancak genler değiştirilmiş hücre tarafından ifade edilebiliyorsa başarılı olur. Konjugasyonda genler, plazmid üzerindeki promotörlerin kontrolü altında bir plazmid üzerinde yer alır. Çıplak DNA'nın hücreye girdiği transformasyon ve transdüksiyonda, DNA genetik ifade oluşmadan hücre tarafından kolayca parçalanabilir. Genlerin ifade edilebilmesi için DNA'nın alıcının kromozomu ile rekombine edilmesi gerekir.
Moleküler rekombinasyonun en yaygın mekanizması, RecA proteinini içeren homolog rekombinasyondur. Bu süreçte iki kaynaktan gelen DNA, bir bölgedeki benzer nükleotid dizilimine dayanarak eşleştirilir. Bir endonükleaz bir ipliği keserek RecA'nın farklı ipliklerden bazları eşleştirmesine izin verir, bu süreç iplik istilası olarak bilinir. DNA molekülleri arasındaki çapraz geçiş, DNA'yı iki ayrı dsDNA molekülü halinde kesen ve yeniden birleştiren resolvaz ile çözülür.
Rekombinasyon, virüslerin genomunu konakçılarının kromozomlarına yerleştirmek için sıklıkla kullandığı, özgül site rekombinasyonunu kullanarak da gerçekleşebilir. Bu tür rekombinasyon aynı zamanda transposable elementler tarafından da kullanılır (bir sonraki bölüme bakınız).
Değiştirilebilir Elemanlar
Son olarak, mikrobiyal genetik konusunu en azından değiştirilebilir elemanların veya "atlayan genlerin" rolünü keşfetmeden bırakmamalıyız. Bunlar bakteriyel genlerin aktivasyonu ve inaktivasyonunda çok büyük bir rol oynayabilirken, en iyi açıklama 1983 yılında araştırmasıyla Nobel Ödülü kazanan Barbara McClintock'un mısırdaki çalışmalarından gelmektedir. Değiştirilebilir elemanların bir organizma içindeki genlerin aktivasyonundan veya inaktivasyonundan sorumlu olabileceğini göstermiştir.
Transpoze edilebilir elemanlar, nispeten basit yapıda olup, transpozisyon olarak bilinen bir süreçle DNA molekülü içinde bir konumdan diğerine geçmek üzere tasarlanmıştır. Tüm değiştirilebilir elemanlar, transpozisyonun gerçekleşmesini sağlayan enzim olan transpozaz enzimini kodlar ve her iki uçta kısa ters çevrilmiş tekrarlara (IR'ler) sahiptir.

En basit transposable eleman, transpozaz ve değişen uzunluklarda IR'ler içeren bir ekleme dizisidir (IS). Bir transpozon tipik olarak ek genler içerir ve tam türü transpozondan transpozona büyük ölçüde değişir. Bir transpozon bir konumdan çıkarılıp başka bir konuma yerleştirilebilir (kes-yapıştır modeli), bu işlem muhafazakar transpozisyon olarak bilinir. Alternatif olarak, replikatif transpozisyon olarak bilinen bir süreçte, kopyanın ikinci bir bölgeye yerleştirilmesiyle çoğaltılabilir.
Önceki Ders: Türüne Göre Sınıflandırma
Sonraki Ders: Genetik Mühendisliği

Yorumlar
Yorum Gönder