Amelojenez
Odontoblast ve ameloblast oluşumunun animasyonlu gösterimi.
İndüksiyon (veya Başlatma)
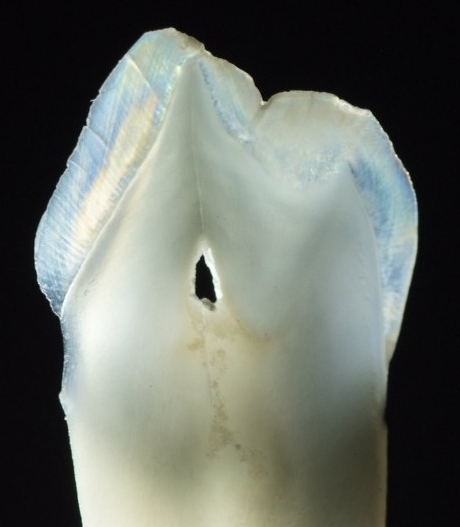
Bu paragraf gözden geçirilmelidir. Amelogenez diş gelişiminin çan aşamasında başlar. Bu aşamada, ektodermal hücreler invajine olarak iç mine epiteli (IEE) ve dış mine epiteli (OEE) olarak adlandırılan iki tek katlı kübik hücre tabakası oluşturur. IEE ve OEE arasında, stratum intermedium ve stellat retikulum hücreleri gibi daha fazla epitel hücresi dağılmıştır. Bu hücreler toplu olarak mine organı olarak bilinir.
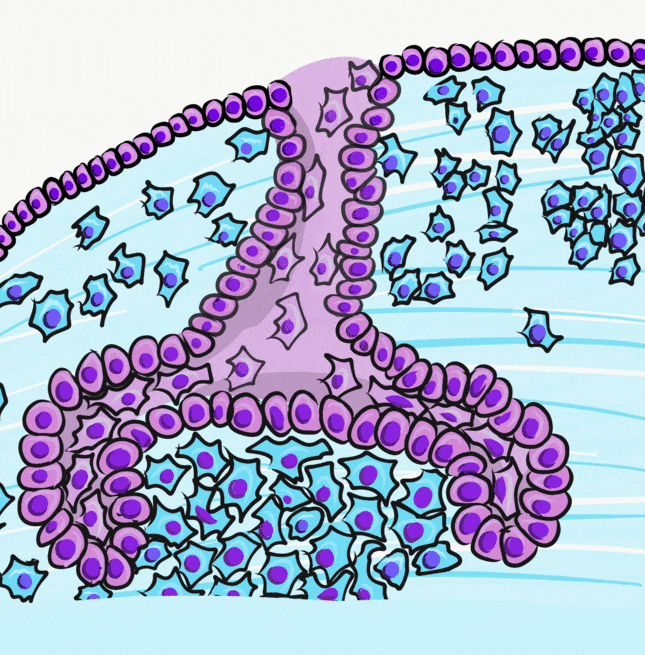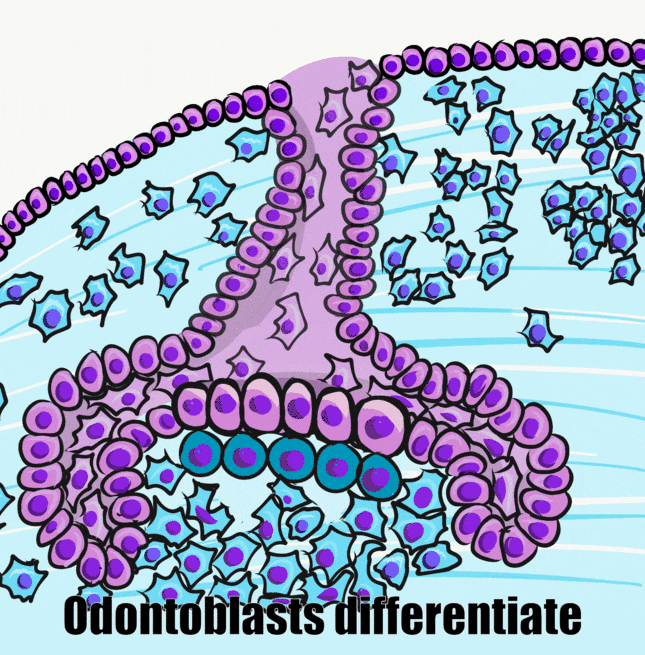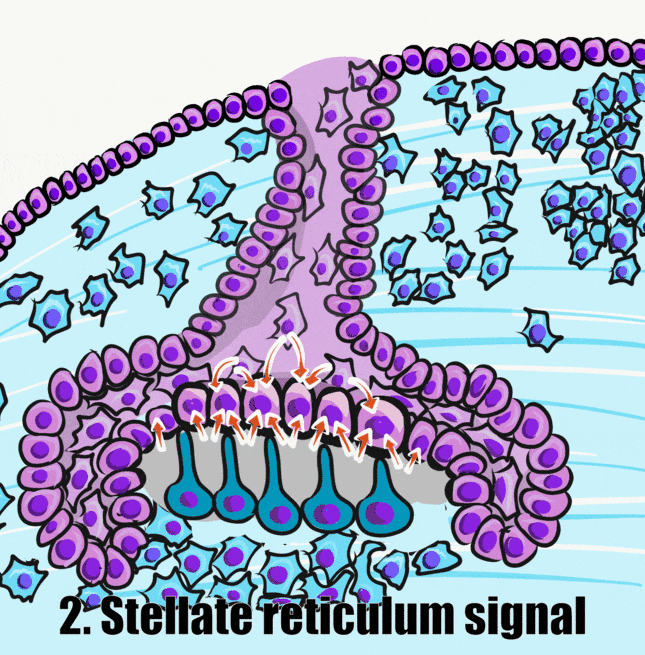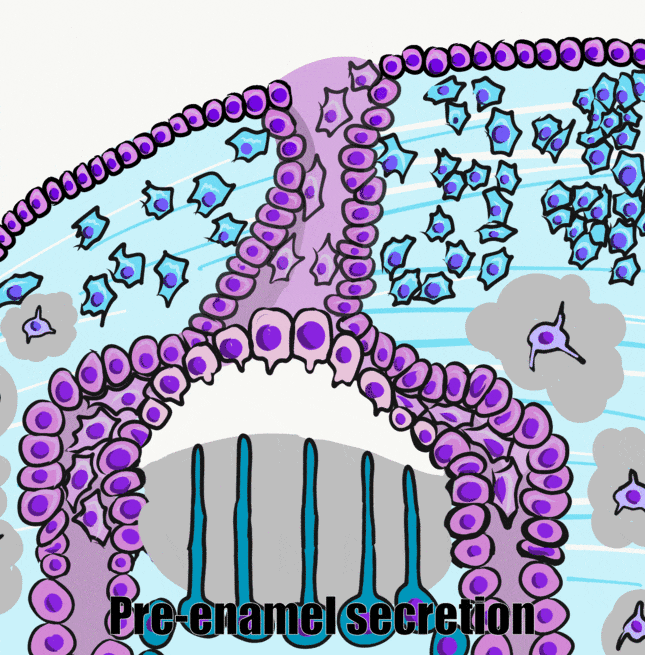
Şimdi bir önceki bölümde öğrendiklerimize ekleme yapalım. Mine organı, dental papilla olarak bilinen bir nöro-mezenkimal kök hücre kümesi etrafında büyür. Başlangıçta, bir bazal membran ikisini ayırır. Farklılaşmanın ilk görünür işareti, dental papillanın yanındaki küboidal IEE hücreleri uzayarak daha sütunlu bir şekle dönüştüğünde ortaya çıkar. Çekirdekleri hücrenin apikal tarafına taşınırken, salgı için gerekli organeller bazal tarafta birikir. Bu uzun hücreleri pre-ameloblast olarak adlandırıyoruz. Morfolojideki bu ilk değişikliği neyin tetiklediği kesin değildir, kanıtlar Wnt morfojen ailesinin bir üyesinin düzlemsel hücre polarite sinyallerini aktive ettiğini göstermektedir. Mitoz geçirme yeteneği durur, yani çan aşamasından sonra yeni pre-ameloblast oluşmaz. Bazal membranın ortadan kalkması, hemi-desmozom bağlantılarının kaybı nedeniyle bu hücrelerin daha kolay hareket etmesini sağlar.
Pre-ameloblastlar, BMP ailesinin bir üyesi de dahil olmak üzere bir dizi kısa menzilli morfojen salgılar (yukarıdaki şekil). Bu, dental papilladaki komşu nöro-mezenkimal kök hücrelerin odontoblastlara farklılaşmasını ve pre-dentin salgılamaya başlamasını sağlar. Bu da pre-ameloblastları pre-dentin ile doğrudan temasa geçirir. Pre-ameloblastlar üzerindeki integrinler, bazal membrandaki tipin (kolajen tip IV) aksine pre-dentinde bulunan bir kolajen formuyla (kolajen tip I) temas eder. Ek olarak, yeni oluşan odontoblastlar BMP'ler de dahil olmak üzere çeşitli morfojenler salgılar. Önemli kavram, ameloblastların farklılaşmasını indükleyen sinyalin yarısının pre-dentin tarafından gelmesidir.
Bu sinyalin ikinci yarısı ters yönden, stellat retikulumdan gelir. Stellat retikulum hücreleri tarafından üretilen böyle bir sinyal FGF ailesinin bir üyesini içerir. Bu, pre-ameloblastlarda transkripsiyon faktörlerinin aktivasyonuna yol açar ve bu da mine salgılanmasında rol oynayan genlerin ifadesini yukarı doğru düzenler. Amelogenezde rol oynayan transkripsiyon faktörleri arasında bir homeobox geni olan MSX-2 ve osteoblastların farklılaşmasında da rol oynayan bir transkripsiyon faktörü olan RUNX2 bulunur. Mikroskop altında görülebilen pre-ameloblastlar tekrar şekil değiştirerek kısalır ve bazal tarafta Tomes prosesi olarak bilinen bir yumru geliştirir. İlgili bazı moleküllerin bir özeti aşağıdaki tabloda yer almaktadır. İsimler, olayların genel sırasından daha az önemlidir: morfojenden reseptöre, transkripsiyon faktöründen gen ifadesine kadar.
Morfojenler | Reseptörler | Transkripsiyon faktörleri | Upregüle genler |
Kolajen (pre-dentin) BMP (odontoblastlar) | İntegrinler BMP-R | MSX-2 DLX1 RUNX2 | Enamelin Amelogenin Matris Metalloproteinaz |
FGF (stellat retikulum) | FGF-R | MSX-2 DLX1 RUNX2 | Enamelin Amelogenin Matris Metalloproteinaz |
Apozisyon
Ameloblastlar morfolojilerini değiştirdikten sonra Tomes prosesinden mine matriksi salgılarlar. Bu ECM, kemik dokusunun ECM'sine benzer, ancak önemli farklılıkları vardır. Kemik matrisi gibi, mine matrisi de ilk olarak çoğunlukla protein ve sudan oluşan daha yumuşak bir formda (pre-enamel) salgılanır ve daha sonra mineralize olur. Mine matrisi Kalsiyum-Fosfat kristalleri ve organik bileşenler içerir. Mine, olgunlaştıktan sonra daha yüksek bir mineral yüzdesine sahip olur ve bu da onu insan vücudundaki en sert madde haline getirir. Ancak kemik dokusunun aksine mine matrisinde çok az kolajen bulunur ya da hiç bulunmaz. Bunun yerine, ameloblastlar amelogeninler ve enamelinler olarak bilinen diğer proteinleri salgılarlar. Bu proteinler, Kalsiyum ve Fosfatın kristaller oluşturmasına yardımcı olan çekirdeklenme merkezleri olarak işlev görür. Bu proteinleri hücrelerden ziyade cansız kimyasallar için iskeleler olarak düşünebiliriz. Burada hatırlanması gereken önemli gerçek, kolajenin genellikle mezodermden farklılaşan fibroblastlar (ve mezenşimden türeyen diğer hücreler) tarafından yapılan bir protein olduğudur. Ameloblast soyu ise ektodermden gelmektedir. Muhtemelen kolajen benzeri bağ dokusu genleri metillenmiş ve histonların etrafına paketlenmiştir ve transkripsiyon için uygun değildir.
Çan aşamasındaki mine organının şekli kuronun şeklini belirler. Yeni ameloblastlar oluşmaz çünkü bunlar pre-ameloblastlara farklılaştıktan sonra mitoz geçirme yeteneklerini kaybederler. Ayrıca, kalın mine ve (olgun) dentin tabakaları ikisini birbirinden ayırdığında hiçbir kök hücre hem pre-dentin hem de stellat retikulum ile temas edemez. Bu nedenle, farklılaşma yoluyla yeni ameloblastlar da oluşmaz. Pre-ameloblastların çan evresinde çoğunlukla ağız boşluğuna bakacak şekilde sıralanması nedeniyle, mine çiğneme yüzeylerinde daha kalın, servikal bölgelerde ise daha incedir.
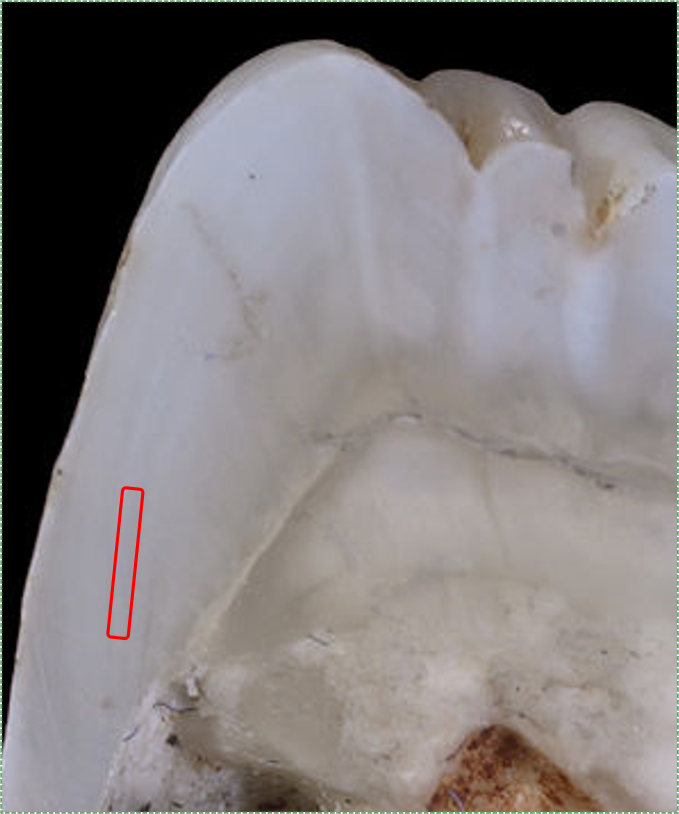
Tomes prosesi ile ameloblastlar tarafından salgılanan mine, eski tabakaların üzerine yeni ön mine tabakaları bırakarak ameloblastları dentinden daha uzağa iter. Ameloblastlar, birçok insan hücresi gibi, tek tip bir hızda çalışmazlar. Yaklaşık her 4-12 günde bir mine birikim oranında bir değişiklik olur. Mine birikiminin hızlanması ve yavaşlamasının bir sonucu olarak, enine kesitte bakıldığında daha açık ve daha koyu mine bantları (daha az yoğun ve daha yoğun) görülebilir. Bu görünür bantlar Retzius Çizgileri (veya Retzius şeritleri, yukarıdaki şekil) olarak bilinir. Bu bantlardan biri olan neonatal çizgi özellikle koyudur çünkü ameloblastlar doğduğumuz gün boyunca istirahat etmişlerdir. Doğumdan sonra, Retzius Çizgilerinin kenarları dişlerin yüzeyinde, özellikle de ön dişlerin lingual yüzeyinde görülebilir, bu durumda perikimata (eski metinlerde Pickerill'in imbrikasyon çizgileri) olarak adlandırılırlar. Zaman içinde kimyasal ve fiziksel aşınma perikimatayı aşındırır.
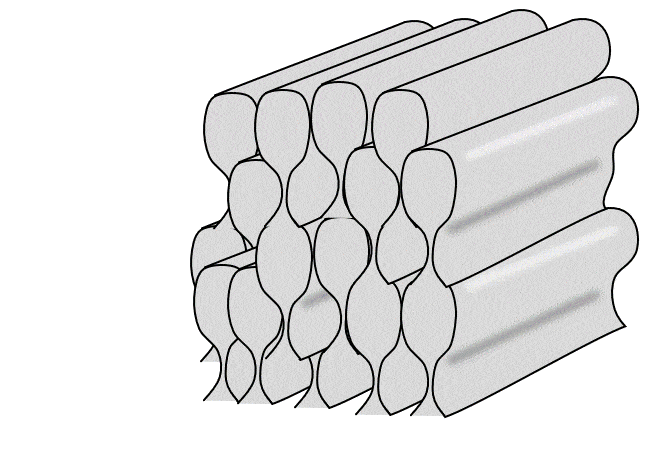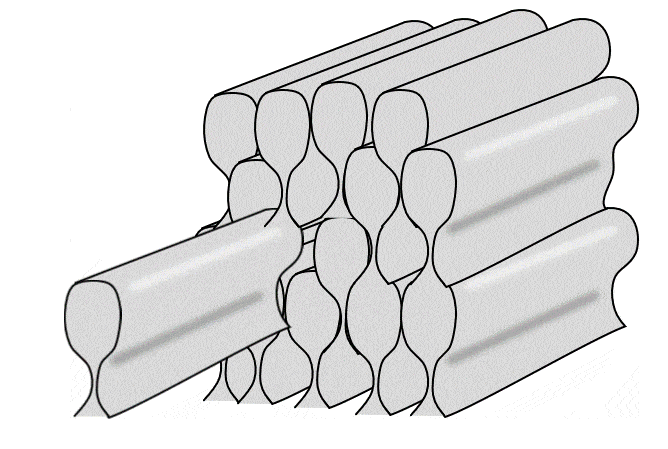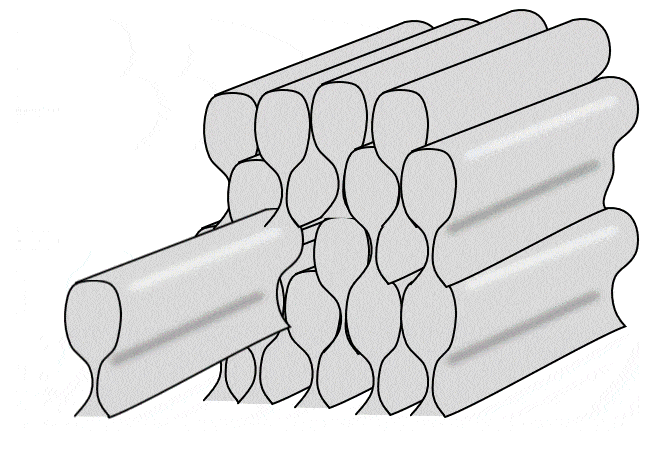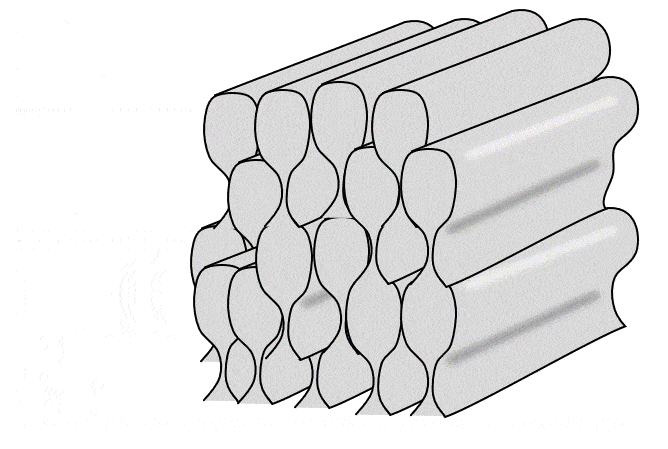
Tüm ameloblastlar bir sıra halinde, eskinin üzerine yeni olmak üzere yatay mine katmanları üretir. Ancak kabaca bir ameloblast dik yönde tek bir mine çubuğu (veya prizma) üretir (bir mine çubuğu yapmak için 4 ameloblast gerekir, ancak her bir ameloblast 4 farklı çubuk üzerinde çalışır). Çubuklar kare değil yuvarlak (neredeyse altıgen) olduğu için aralarında boşluklar vardır. Bu boşluklar boş değildir, çubuklar arası mine adı verilen biraz farklı bir mine formuyla doludur (çubukların içindeki mineye çubuk minesi denir).
İki yukarıdaki şekle yakından bakarsanız, Retzius ve DEJ çizgilerine dik uzanan çizgiler olan Hunter-Schreger bantlarını (HSB) fark edebilirsiniz. Her çizgi küçük bir ameloblast grubunun katkılarını temsil etmektedir. Görmesi biraz zorsa, bunun nedeni her bir Hunter-Schreger bandının milimetrenin yalnızca 1/10'u kalınlığında olmasıdır. Aradığınız şey, bir ders kitabının ön kenarına benzer bir desendir (grup olarak tüm sayfalar). Ameloblastlar düz çizgiler halinde çalışmaz, bantlar hafifçe kıvrılır - suya düşürdüğünüz ve kitapçıya geri satamadığınız ders kitabının sayfaları gibi. Ayrıca, farklı bölgeler farklı eğri açılarına ve desenlerine sahiptir. Bu kıvrımlar tesadüfi değildir, minenin gerilme mukavemetini arttırırlar. Daha önce odun kestiyseniz, baltayı damar boyunca indirdiğinizde bunu yapmanın kolay olduğunu bilirsiniz. Açılı vurursanız, size zarar verebilir. Hunter-Schreger bantlarının kıvrımlı olması, bir dişin kolayca kesilebileceği tek bir damar olmamasını sağlar. Minede çatlaklar oluşur, ancak çatlaklar genellikle yarı yolda durur. Çubuk eğriliğine rehberlik eden düzlemsel hücre polarite sinyalleri tanımlanmıştır (Wnt morfojenlerinin antagonistleri). Sonuç olarak mine, cam kırılganlığına sahip olmasına rağmen insan vücudundaki en dirençli dokudur. Dentin, mineden farklı olarak özel isimler alan benzer kıvrımlara (çoğul) sahiptir. Eğlenceli bir yan not olarak ya da yerel hayvanat bahçenizde veteriner diş hekimi olarak kariyer yapmayı planlayanlarla ilgili olarak, sırtlanların Hunter-Schreger bantları zig-zag şeklinde oluşur ve minelerinin gerilme mukavemetini daha da artırarak kemikleri çiğnemelerine olanak tanır.
Olgunlaşma
Mine ilk olarak pre-enamel adı verilen protein açısından zengin yapışkan bir madde olarak salgılanır. Minenin iki ana organik bileşeni amelogenin ve enamelin proteinleridir. Ameloblastlar ayrıca bir sindirim enzimi de salgılar. Bir matriks metalloproteinaz olan bu enzim, kolajeni sindirerek ameloblastların üstteki mezenşimin zemin maddesi boyunca göç etmesini kolaylaştırır. Bu gereklidir çünkü ameloblastlar Tomes prosesinden gelen ön mineyi biriktirirken hücreleri dışarı doğru iter. Hatırlarsanız, matriks metalloproteinazlar aynı zamanda damak oluşumu sırasında dokuların düzgün bir şekilde göç etmesinde ve dişlerin sürmesinde de rol oynarlar. Matriks metalloproteinazlar, ECM'ye salgılandıktan sonra ameloblastlar tarafından yapılan mine proteinlerini işleyerek ikinci bir rol oynarlar. Enzimler amelogeninleri ve enamelinleri modifiye ederek (sindirmeyerek) kalsiyum ve fosfatın kristalleşmesini katalize etme yeteneklerini geliştirir. Bu, ön minenin salgılanması ile mineralizasyonu arasında bir gecikme yaratır. Gecikme olmazsa, ameloblastlar kendi ECM'leri tarafından tuzağa düşürülebilir ve amelogenezi durdurabilir.
Dentin ve sementumun aksine mine, mineralizasyondan sonra önemli miktarda yeniden şekillenmeye uğrar. Ön mine oluştuktan ve mineralize olduktan sonra, ameloblastlar görev değiştirir ve sert minenin olgunlaşarak daha da sert mineye dönüşmesini sağlar. Proteinlerin çoğu uzaklaştırılır, yerini kristalleşen sıvılar alır. Amelogenin ve enamelin endositoz ve protein sindiren enzimlerin salgılanması ile uzaklaştırılır. Bu enzimler arasında matriks metalloproteinaz da yer alır; amelogenin ve enameline yapılan ilk müdahaleler işlevlerini iyileştirir, ancak daha fazla müdahale onları parçalar. Proteinin uzaklaştırılması minenin nispi mineral içeriğini artırarak kemik, dentin ve sementuma kıyasla sertliğini artırır. Bu nedenle ameloblastlar tarafından salgılanan proteini, minenin günde yaklaşık 4 μM hızla üretilmesini sağlayan bir iskele olarak düşünebiliriz. Buna karşılık, diş minesinin florür bazlı yeniden mineralizasyonu küçük bir cetvel kullanılarak ölçülmez, bu da yeniden mineralizasyon hızının mikroskop altında görülemeyecek kadar yavaş olduğunu gösterir.
Bir dizi farklı elektrolit ve molekül, olgunlaşma aşamasında mineye küçük miktarlarda dahil olur. Önemli elektrolitlerden biri florürdür (F-). Florür iyonları negatif yüklüdür ve az sayıda hidroksit (OH-) iyonunun yerini alarak kalsiyum hidroksiapatit kristallerine dahil olur. Bunun önemli klinik etkileri vardır. İlk olarak, az miktarda florür kalsiyum hidroksiapatit kristallerinin gücünü artırır. İkinci olarak, tükürükte ya da minede bulunan florür iyonları bakteriler tarafından salınan asitleri nötralize ederek minenin demineralizasyonunu azaltır ve çürükleri önler. Bununla birlikte, aşırı florür seviyeleri, amelogenezin olgunlaşma aşamasında ameloblastların aktivitesini etkileyerek protein iskelelerini kaldırmalarını azaltır. Bu durum diş yüzeyinde beyaz lezyonlar şeklinde görsel bir değişiklik yaratabilir. Bu lezyonlar, diş minesinin daha yüksek protein ve daha düşük kristal içeriğine sahip olduğu, diş florozisi olarak bilinen lezyonlardır. Her kimyasalda olduğu gibi, zehirli olması dozuna bağlıdır. Diş macunu veya jellerle yapılan topikal uygulama, florürün minenin yüzey katmanlarına dahil olmasını sağlar ve bunun önemli faydaları vardır. En büyük potansiyel fayda, florür maruziyetinin embriyonik gelişimin erken dönemlerinde, amelogenezin appozisyon ve olgunlaşma aşamalarında gerçekleşmesidir.
Önceki Ders: Mine Gelişimine Genel Bakış
Sonraki Ders: Mine Gelişimi Klinik Değerlendirmeler

Yorumlar
Yorum Gönder