Duyusal Algılama
Duyu reseptörlerinin önemli bir rolü, çevremizdeki ortam veya iç ortamımızın durumu hakkında bilgi edinmemize yardımcı olmaktır. Farklı kaynaklardan ve farklı türlerden gelen uyaranlar alınır ve sinir sisteminin elektrokimyasal sinyallerine dönüştürülür. Bu, bir uyaran duyusal bir nöronun hücre zarı potansiyelini değiştirdiğinde meydana gelir. Uyaran, duyu hücresinin merkezi sinir sistemine (MSS) iletilen bir aksiyon potansiyeli üretmesine neden olur ve burada diğer duyusal bilgilerle veya bazen daha yüksek bilişsel işlevlerle entegre edilerek bu uyaranın bilinçli bir algısı haline gelir. Merkezi entegrasyon daha sonra bir motor tepkiye yol açabilir.
Duyusal işlevi duyum ya da algı terimleriyle tanımlamak bilinçli bir ayrımdır. Duyum, uyaran düzeyinde duyusal reseptör hücrelerinin aktivasyonudur. Algı, duyusal uyaranların merkezi olarak işlenerek anlamlı bir örüntüye dönüştürülmesidir. Algı duyuma bağlıdır, ancak tüm duyumlar algılanmaz. Reseptörler, duyumları algılayan hücreler veya yapılardır. Bir reseptör hücresi bir uyaran tarafından doğrudan değiştirilir. Bir transmembran protein reseptörü, çoğunlukla iyon kanallarının açılması veya hücre sinyalizasyon süreçlerindeki değişiklikler yoluyla bir nöronda fizyolojik bir değişikliğe aracılık eden hücre zarındaki bir proteindir. Transmembran reseptörleri ligand adı verilen kimyasallar tarafından aktive edilir. Örneğin, yiyeceklerdeki bir molekül tat reseptörleri için bir ligand görevi görebilir. Tam olarak reseptör olarak adlandırılmayan diğer transmembran proteinleri mekanik veya termal değişikliklere duyarlıdır. Bu proteinlerdeki fiziksel değişiklikler zar boyunca iyon akışını artırır ve duyusal nöronlarda bir aksiyon potansiyeli veya kademeli bir potansiyel oluşturabilir.
Duyusal Reseptörler
Çevredeki uyaranlar, periferik sinir sistemindeki özelleşmiş reseptör hücrelerini harekete geçirir. Farklı türdeki uyaranlar farklı türdeki reseptör hücreleri tarafından algılanır. Reseptör hücreleri üç farklı kriter temelinde tiplere ayrılabilir: hücre tipi, pozisyon ve fonksiyon. Reseptörler, hücre tipi ve algıladıkları uyaranlara göre konumları temelinde yapısal olarak sınıflandırılabilir. Ayrıca, uyaranların iletimi veya mekanik uyaranın, ışığın veya kimyasalın hücre zarı potansiyelini nasıl değiştirdiği temelinde işlevsel olarak da sınıflandırılabilirler.
Yapısal Reseptör Tipleri
Çevre hakkındaki bilgileri yorumlayan hücreler ya (1) serbest bir sinir ucuna sahip olan ve dendritleri bir duyu alacak dokuya gömülü olan bir nöron; (2) duyusal sinir uçlarının duyarlılıklarını artıran bağ dokusu içinde kapsüllendiği kapsüllü bir uca sahip olan bir nöron; ya da (3) belirli bir uyaran türünü yorumlayan farklı yapısal bileşenlere sahip olan özelleşmiş bir reseptör hücresi olabilir (aşağıdaki şekil). Derinin dermisindeki ağrı ve sıcaklık reseptörleri, serbest sinir uçlarına sahip nöronlara örnektir. Ayrıca derinin dermisinde, basınç ve dokunmaya tepki veren kapsüllü sinir uçlarına sahip nöronlar olan lamelli korpüsküller bulunur. Retinadaki ışık uyaranlarına yanıt veren hücreler, özelleşmiş bir reseptör olan fotoreseptörün bir örneğidir.

Reseptörlerin sınıflandırılabilmesinin bir başka yolu da uyaranlara göre konumlarına dayanmaktadır. Eksteroseptör, deride bulunan somatosensör reseptörler gibi dış ortamdaki bir uyaranın yakınında bulunan bir reseptördür. Bir interoseptör, aort veya karotis sinüste kan basıncındaki artışı algılayan reseptörler gibi iç organ ve dokulardan gelen uyarıları yorumlayan bir reseptördür. Son olarak, bir proprioseptör, kas gibi vücudun hareketli bir kısmının yakınında bulunan ve hareket ettikçe dokuların pozisyonlarını yorumlayan bir reseptördür.
Fonksiyonel Reseptör Tipleri
Reseptörlerin üçüncü bir sınıflandırması, reseptörün uyaranları zar potansiyeli değişikliklerine nasıl dönüştürdüğüne göredir. Uyaranlar üç genel tiptedir. Bazı uyaranlar iyonlar ve makromoleküllerdir ve bu kimyasallar hücre zarı boyunca yayıldığında transmembran reseptör proteinlerini etkiler. Bazı uyaranlar, reseptör hücre zarı potansiyellerini etkileyen ortamdaki fiziksel değişikliklerdir. Diğer uyaranlar arasında görünür ışıktan gelen elektromanyetik radyasyon yer alır. İnsanlar için, gözlerimiz tarafından algılanan tek elektromanyetik enerji görünür ışıktır. Yılanların ısı sensörleri, arıların ultraviyole ışık sensörleri veya göçmen kuşlardaki manyetik reseptörler gibi diğer bazı organizmalarda insanlarda bulunmayan reseptörler vardır.
Reseptör hücreleri, ilettikleri uyaranların türüne göre daha da fazla sınıflandırılabilir. Kimyasal uyaranlar, bir nesnenin tadı veya kokusu gibi kimyasal uyaranları yorumlayan bir kemoreseptör tarafından yorumlanabilir. Osmoreseptörler vücut sıvılarının solüt konsantrasyonlarına yanıt verir. Ek olarak, ağrı öncelikle doku hasarından veya benzer yoğun uyaranlardan kaynaklanan kimyasalların varlığını bir nosiseptör aracılığıyla yorumlayan kimyasal bir duyudur. Basınç ve titreşim gibi fiziksel uyaranların yanı sıra ses ve vücut pozisyonu (denge) hissi de bir mekanoreseptör aracılığıyla yorumlanır. Kendine has bir reseptörü olan bir diğer fiziksel uyaran da sıcaklıktır; sıcaklık, normal vücut sıcaklığının üzerindeki (sıcak) veya altındaki (soğuk) sıcaklıklara duyarlı olan bir termoreseptör aracılığıyla algılanır.
Duyusal Modaliteler
Herhangi birine duyuların ne olduğunu sorduğunuzda, muhtemelen beş ana duyuyu (tat, koku, dokunma, işitme ve görme) sıralayacaktır. Ancak bunlar duyuların tamamı değildir. Bu listedeki en belirgin eksiklik dengedir. Ayrıca, basitçe dokunma olarak adlandırılan şey, bu dokunma hislerini algılayan mekanoreseptörlerin türüne göre basınç, titreşim, gerilme ve saç folikülü pozisyonu olarak alt bölümlere ayrılabilir. Göz ardı edilen diğer duyular arasında termoreseptörler tarafından sıcaklık algısı ve nosiseptörler tarafından ağrı algısı yer almaktadır.
Fizyoloji alanında, duyular genel ya da özel olarak sınıflandırılabilir. Genel duyu, vücudun her tarafına dağılmış olan ve diğer organların yapıları içinde reseptör hücreleri bulunan duyudur. Derideki, kaslardaki veya kan damarlarının duvarlarındaki mekanoreseptörler bu türün örnekleridir. Genel duyular genellikle yukarıda açıklandığı gibi dokunma duyusuna veya propriyosepsiyona (vücut hareketi) ve kinesteziye (vücut hareketi) veya otonomik işlevler için en önemli olan viseral bir duyuya katkıda bulunur. Özel bir duyu, göz, iç kulak, dil veya burun gibi kendisine ayrılmış belirli bir organı olan duyudur.
Duyuların her biri duyusal modalite olarak adlandırılır. Modalite, transdüksiyon fikrine benzer şekilde bilginin kodlanma biçimini ifade eder. Ana duyusal modaliteler, her birinin nasıl iletildiği temelinde tanımlanabilir. Kimyasal duyular tat ve kokudur. Genellikle dokunma olarak adlandırılan genel duyu, nosisepsiyon veya ağrı şeklindeki kimyasal hissi içerir. Basınç, titreşim, kas gerilmesi ve saçın harici bir uyaran tarafından hareket ettirilmesi, mekanoreseptörler tarafından algılanır. İşitme ve denge de mekanoreseptörler tarafından algılanır. Son olarak, görme fotoreseptörlerin aktivasyonunu içerir.
Sayıları 17'ye kadar çıkabilen tüm farklı duyusal modaliteleri listelemek, beş ana duyuyu daha büyük duyunun daha spesifik kategorilerine veya alt modalitelerine ayırmayı içerir. Bireysel bir duyusal modalite, belirli bir uyaran türünün duyumunu temsil eder. Örneğin, somatosensasyon olarak bilinen genel dokunma duyusu, hafif basınç, derin basınç, titreşim, kaşıntı, ağrı, sıcaklık veya saç hareketi olarak ayrılabilir.
Gustation (Tat)
Tat alma duyusu veya gustasyon içinde sadece birkaç tanınmış alt modalite mevcuttur. Yakın zamana kadar sadece dört tat tanınıyordu: tatlı, tuzlu, ekşi ve acı. Yirminci yüzyılın başlarında yapılan araştırmalar, 1980'lerin ortalarında beşinci tat olan umaminin tanınmasına yol açtı. Umami, "lezzetli tat" anlamına gelen Japonca bir kelimedir ve genellikle tuzlu anlamına gelecek şekilde çevrilir. Çok yeni araştırmalar, yağlar veya lipitler için de altıncı bir tat olabileceğini öne sürmüştür.
Gustasyon, dil ile ilişkili özel bir duyudur. Dilin yüzeyi, ağız boşluğunun geri kalanıyla birlikte, çok katlı bir yassı epitel ile kaplıdır. Papilla adı verilen kabarık çıkıntılar tat alma iletimini sağlayan yapıları içerir. Görünüşlerine göre dört tip papilla vardır (aşağıdaki şekil): dairesel, yapraksı, filiform ve fungiform. Papillaların yapısında, tat uyaranlarının iletimi için özelleşmiş tat alma reseptör hücreleri içeren tat tomurcukları bulunur. Bu reseptör hücreleri, alınan gıdaların içerdiği kimyasallara karşı duyarlıdır ve gıdadaki kimyasal miktarına bağlı olarak nörotransmitter salgılarlar. Tat hücrelerinden gelen nörotransmitterler yüz, glossofarengeal ve vagus kraniyal sinirlerindeki duyusal nöronları aktive edebilir.

Tuzlu tat basitçe tükürükteki sodyum iyonlarının (Na+) algılanmasıdır. Tuzlu bir şey yediğinizde, tuz kristalleri Na+ ve Cl- bileşen iyonlarına ayrışır ve bunlar ağzınızdaki tükürükte çözülür. Na+ konsantrasyonu tat alma hücrelerinin dışında yüksek hale gelir ve iyonun hücrelere difüzyonunu sağlayan güçlü bir konsantrasyon gradyanı oluşturur. Bu hücrelere Na+ girişi, hücre zarının depolarizasyonuna ve bir reseptör potansiyelinin oluşmasına neden olur.
Ekşi tat, H+ konsantrasyonunun algılanmasıdır. Tıpkı tuzlu tatlardaki sodyum iyonlarında olduğu gibi, bu hidrojen iyonları hücreye girer ve depolarizasyonu tetikler. Ekşi tatlar, esasen, yiyeceklerimizdeki asitlerin algılanmasıdır. Tükürükteki hidrojen iyonu konsantrasyonlarının artması (tükürük pH'ının düşmesi) tat alma hücrelerinde giderek daha güçlü kademeli potansiyelleri tetikler. Örneğin, sitrik asit içeren portakal suyunun tadı ekşidir çünkü pH değeri yaklaşık 3'tür. Tabii ki, ekşi tadın maskelenmesi için genellikle tatlandırılır.
İlk iki tat (tuzlu ve ekşi) Na+ ve H+ katyonları tarafından tetiklenir. Diğer tatlar, gıda moleküllerinin G proteinine bağlı bir reseptöre bağlanmasından kaynaklanır. Bir G proteini sinyal iletim sistemi sonuçta tat alma hücresinin depolarizasyonuna yol açar. Tatlı tat, tat alma hücrelerinin tükürükte çözünmüş glikoz varlığına karşı duyarlılığıdır. Fruktoz gibi diğer monosakkaritler veya aspartam (NutraSweet™), sakarin veya sukraloz (Splenda™) gibi yapay tatlandırıcılar da tatlı reseptörlerini aktive eder. Bu moleküllerin her biri için afinite değişir ve bazıları G proteinine bağlı reseptöre farklı şekilde bağlandıkları için glikozdan daha tatlı bir tada sahip olurlar.
Acı tat, gıda moleküllerinin G proteinine bağlı reseptörlere bağlanması bakımından tatlıya benzer. Ancak, acı tada sahip çok çeşitli moleküller olduğu için bunun gerçekleşebileceği bir dizi farklı yol vardır. Bazı acı moleküller tat alma hücrelerini depolarize ederken, diğerleri tat alma hücrelerini hiperpolarize eder. Benzer şekilde, bazı acı moleküller tat alma hücrelerindeki G proteini aktivasyonunu artırırken, diğer acı moleküller G proteini aktivasyonunu azaltır. Spesifik yanıt, reseptöre hangi molekülün bağlandığına bağlıdır.
Acı tat veren moleküllerin önemli bir grubu alkaloidlerdir. Alkaloidler, kahve, şerbetçiotu (birada), tanen (şarapta), çay ve aspirin gibi acı tat veren bitkisel ürünlerde yaygın olarak bulunan azot içeren moleküllerdir. Toksik alkaloidler içerdiğinden, bitki mikrop enfeksiyonuna karşı daha az duyarlı ve otçullar için daha az çekicidir.
Bu nedenle, acı tadın işlevi öncelikle zehirlerin yutulmasını önlemek için öğürme refleksini uyarmakla ilgili olabilir. Bu nedenle, normalde yenen birçok acı gıda, daha lezzetli hale getirmek için genellikle tatlı bir bileşenle birleştirilir (örneğin kahvede krema ve şeker). Acı reseptörlerinin en yüksek konsantrasyonunun, öğürme refleksinin zehirli yiyecekleri tükürmeye devam edebileceği arka dilde olduğu görülmektedir.
Umami olarak bilinen tat genellikle tuzlu tat olarak adlandırılır. Tatlı ve acı gibi, G proteinine bağlı reseptörlerin belirli bir molekül tarafından etkinleştirilmesine dayanır. Bu reseptörü aktive eden molekül L-glutamat amino asididir. Bu nedenle, umami tadı genellikle protein açısından zengin gıdalar tüketilirken algılanır. Şaşırtıcı olmayan bir şekilde, et içeren yemekler genellikle tuzlu olarak tanımlanır.
Tat hücreleri tat molekülleri tarafından aktive edildikten sonra, duyusal nöronların dendritleri üzerine nörotransmitterler salgılarlar. Bu nöronlar yüz ve glossofaringeal kraniyal sinirlerin bir parçasıdır ve vagus siniri içinde öğürme refleksine adanmış bir bileşendir. Yüz siniri dilin ön üçte birlik kısmındaki tat tomurcuklarına bağlanır. Glossofarengeal sinir, dilin arka üçte ikisindeki tat tomurcuklarına bağlanır. Vagus siniri, acı gibi zararlı uyaranlara karşı daha hassas olan dilin en arka kısmındaki tat tomurcuklarına bağlanır.
| İNTERAKTİF BAĞLANTI Philadelphia, Pennsylvania'daki Monell Kimyasal Duyu Merkezi'nden Dr. Danielle Reed hakkında bilgi edinmek için bu videoyu izleyin; kendisi duyusal deneyimleri sayesinde erken yaşta bilime ilgi duymaya başlamıştır. Tanıdığı diğer insanlarla karşılaştırıldığında kendi tat alma duyusunun benzersiz olduğunu fark etti. Şimdi, insanlar arasındaki genetik farklılıkları ve tat uyaranlarına karşı duyarlılıklarını inceliyor. Videoda, renkli bir boya ile kaplanmış olan dilini dışarı çıkaran bir kişinin kısa bir görüntüsü yer alıyor. Dr. Reed dil yüzeyindeki papillaları bu şekilde görselleştirip sayabilmektedir. İnsanlar dillerindeki papillaların yoğunluğuna göre "tat alan" ve "tat almayan" olarak bilinen iki gruba ayrılır ve bu da tat tomurcuklarının sayısını gösterir. Tat almayanlar yiyeceklerin tadına bakabilir, ancak acı gibi belirli tatlara karşı o kadar hassas değildirler. Dr. Reed kendisinin tat almayan biri olduğunu keşfetti, bu da acıyı neden tanıdığı diğer insanlardan farklı algıladığını açıklıyor. Tatlara karşı çok hassas mısınız? Ailenizin üyeleri arasında herhangi bir benzerlik görebiliyor musunuz? |
Olfaction (Koku)
Tat alma duyusu gibi koku alma duyusu da kimyasal uyaranlara karşı duyarlıdır. Koku alma reseptör nöronları üst burun boşluğu içinde küçük bir bölgede bulunur (aşağıdaki şekil). Bu bölge koku epiteli olarak adlandırılır ve bipolar duyusal nöronlar içerir. Her koku alma duyu nöronunun, epitelin apikal yüzeyinden boşluğu kaplayan mukusa uzanan dendritleri vardır. Havadaki moleküller burun yoluyla solunduğunda, koku epitel bölgesinin üzerinden geçer ve mukus içinde çözülür. Bu koku molekülleri, onları mukus içinde çözünmüş halde tutan proteinlere bağlanır ve koku alma dendritlerine taşınmalarına yardımcı olur. Koku verici-protein kompleksi, bir koku alma dendritinin hücre zarı içindeki bir reseptör proteine bağlanır. Bu reseptörler G proteinine bağlıdır ve koku alma nöronlarında kademeli bir zar potansiyeli üretecektir.
Bir koku alma nöronunun aksonu epitelin bazal yüzeyinden, etmoid kemiğin kribriform plakasındaki bir koku alma forameninden geçerek beyne uzanır. Koku alma yolu olarak adlandırılan akson grubu, ön lobun ventral yüzeyindeki koku alma ampulüne bağlanır. Aksonlar buradan çeşitli beyin bölgelerine gitmek üzere ayrılır. Bazıları serebruma, özellikle de temporal lobun inferior ve medial bölgelerinde bulunan primer koku korteksine gider. Diğerleri ise limbik sistem ve hipotalamus içindeki yapılara yansır ve burada kokular uzun süreli hafıza ve duygusal tepkilerle ilişkilendirilir. Kişinin doğduğu yerle ilişkilendirilen yemek kokusu gibi bazı kokular duygusal anıları bu şekilde tetikler. Koku, serebral kortekse bağlanmadan önce talamusta sinaps yapmayan tek duyusal modalitedir. Koku alma sistemi ile serebral korteks arasındaki bu yakın bağlantı, kokunun anıların ve duyguların güçlü bir tetikleyicisi olabilmesinin bir nedenidir.
Koku alma hücreleri de dahil olmak üzere burun epiteli havadaki toksik kimyasallardan zarar görebilir. Bu nedenle, koku alma nöronları burun epiteli içinde düzenli olarak değiştirilir, ardından yeni nöronların aksonları koku alma ampulünde uygun bağlantılarını bulmalıdır. Bu yeni aksonlar, kraniyal sinirde zaten mevcut olan aksonlar boyunca büyür.

| …BOZUKLUKLARI Koku Sistemi: Anosmi Birçok araba kazasında olduğu gibi yüze gelen künt kuvvet travması, koku alma sinirinin kaybına ve ardından koku alma duyusunun kaybına yol açabilir. Bu durum anosmi olarak bilinir. Beynin ön lobu etmoid kemiğe göre hareket ettiğinde, koku alma yolu aksonları birbirinden ayrılabilir. Profesyonel dövüşçüler, yüz ve kafaya tekrarlanan travmalar nedeniyle sıklıkla anosmi yaşarlar. Buna ek olarak, antibiyotikler gibi bazı farmasötikler, tüm koku alma nöronlarını bir anda öldürerek anosmiye neden olabilir. Koku alma sinirinde hiç akson yoksa, yeni oluşan koku alma nöronlarından gelen aksonların onları koku alma ampulü içindeki bağlantılarına götürecek hiçbir kılavuzu yoktur. Solunum yolu enfeksiyonları veya alerjilere bağlı enflamatuar tepkilerden kaynaklananlar gibi geçici anosmi nedenleri de vardır. Koku alma duyusunun kaybı yiyeceklerin tadının yavan olmasına neden olabilir. Koku alma duyusu bozulmuş bir kişi, yiyeceklerin tadına varabilmek için ek baharat ve çeşni katmaya ihtiyaç duyabilir. Anosmi bazı hafif depresyon tablolarıyla da ilişkili olabilir, çünkü yemekten alınan zevkin kaybı genel bir umutsuzluk hissine yol açabilir. Koku alma nöronlarının kendilerini yenileme yeteneği yaşla birlikte azalır ve bu da yaşa bağlı anosmiye yol açar. Bu durum bazı yaşlıların neden gençlere kıyasla yiyeceklerini daha fazla tuzladıklarını açıklamaktadır. Ancak bu artan sodyum alımı kan hacmini ve kan basıncını artırarak yaşlılarda kardiyovasküler hastalık riskini artırabilir. |
Audition (İşitme)
İşitme veya duyma, ses dalgalarının kulak yapıları tarafından mümkün kılınan sinirsel bir sinyale dönüştürülmesidir (aşağıdaki şekil). Başın yan tarafındaki büyük, etli yapı kulak kepçesi olarak bilinir. Kulak kepçesinin C şeklindeki kıvrımları ses dalgalarını işitme kanalına doğru yönlendirir. Kanal, kafatasına temporal kemiğin dış kulak yolundan girer. İşitme kanalının sonunda, ses dalgaları tarafından vurulduktan sonra titreşen timpanik zar veya kulak zarı bulunur. Kulak kepçesi, kulak kanalı ve timpanik zar genellikle dış kulak olarak adlandırılır. Orta kulak, kemikçik adı verilen üç küçük kemiğin kapladığı bir boşluktan oluşur. Bu üç kemikçik malleus, incus ve stapes olup Latince isimleri kabaca çekiç, örs ve üzengi anlamına gelmektedir. Malleus timpanik zara bağlıdır ve incus ile eklemleşir. İnkus da stapes ile eklemleşir. Stapes daha sonra ses dalgalarının sinirsel bir sinyale dönüştürüleceği iç kulağa bağlanır. Orta kulak, timpanik zar boyunca hava basıncını dengelemeye yardımcı olan östaki borusu aracılığıyla yutağa bağlanır. Tüp normalde kapalıdır ancak yutkunma veya esneme sırasında yutak kasları kasıldığında açılır.
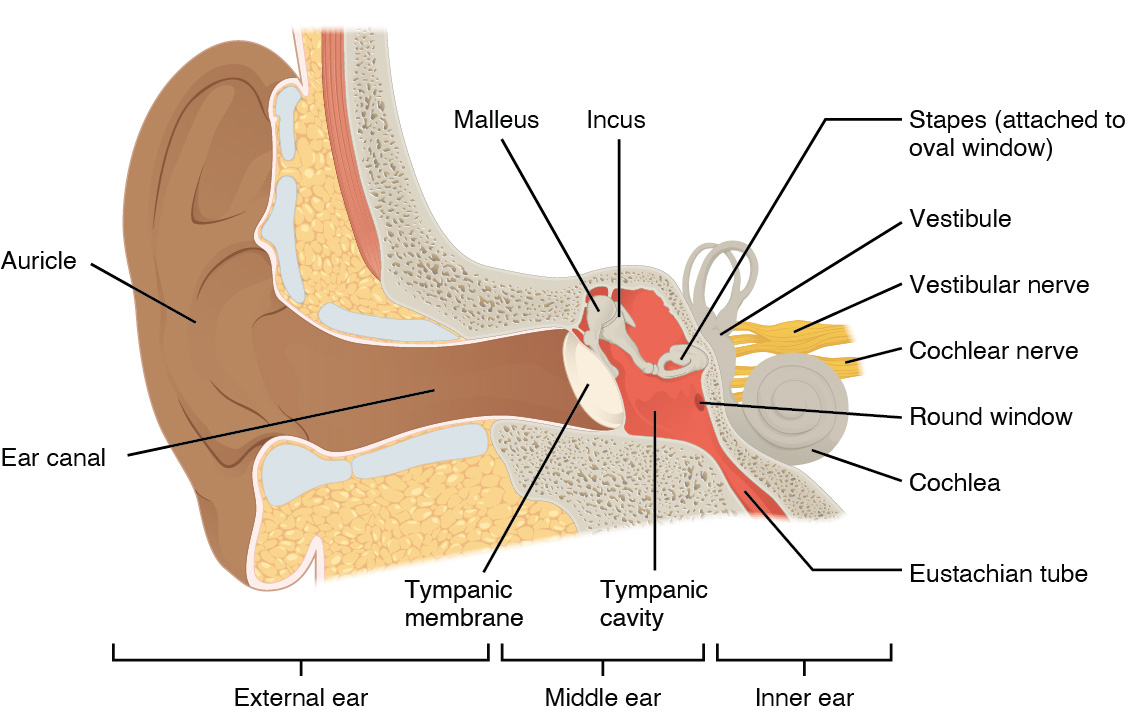
İç kulak, temporal kemiğin içine gömülü bir dizi kanaldan oluştuğu için genellikle kemikli bir labirent olarak tanımlanır. Koklea ve vestibül olmak üzere sırasıyla işitme ve dengeden sorumlu iki ayrı bölgeye sahiptir. Bu iki bölgeden gelen sinirsel sinyaller ayrı lif demetleri aracılığıyla beyin sapına iletilir. Ancak bu iki farklı demet, vestibülokoklear sinir olarak iç kulaktan beyin sapına birlikte gider. Ses, spiral gangliyonların duyusal nöronlarını içeren iç kulağın koklear bölgesinde sinir sinyallerine dönüştürülür. Bu gangliyonlar iç kulağın spiral şeklindeki kokleası içinde yer alır. Koklea, oval pencere aracılığıyla stapes'e bağlanır.
Oval pencere, koklea içinde skala vestibuli adı verilen sıvı dolu bir tüpün başlangıcında yer alır. Skala vestibuli, oval pencereden uzanarak kokleanın ses ileten nöronları içeren merkezi boşluğu olan koklear kanalın üzerinde ilerler. Kokleanın en üst ucunda, skala vestibuli koklear kanalın üstünden kıvrılır. Artık skala timpani olarak adlandırılan sıvı dolu tüp, bu kez koklear kanalın altından geçerek kokleanın tabanına geri döner. Skala timpani, skala içindeki sıvıyı içeren bir zarla kaplı olan yuvarlak pencerede sonlanır. Kemikçiklerin titreşimleri oval pencereden geçerken, skala vestibuli ve skala timpani sıvısı dalga benzeri bir hareketle hareket eder. Sıvı dalgalarının frekansı ses dalgalarının frekanslarıyla eşleşir (aşağıdaki şekil). Yuvarlak pencereyi örten zar, sıvının skala timpani içindeki hareketiyle dışarı doğru şişecek veya içeri doğru büzüşecektir.

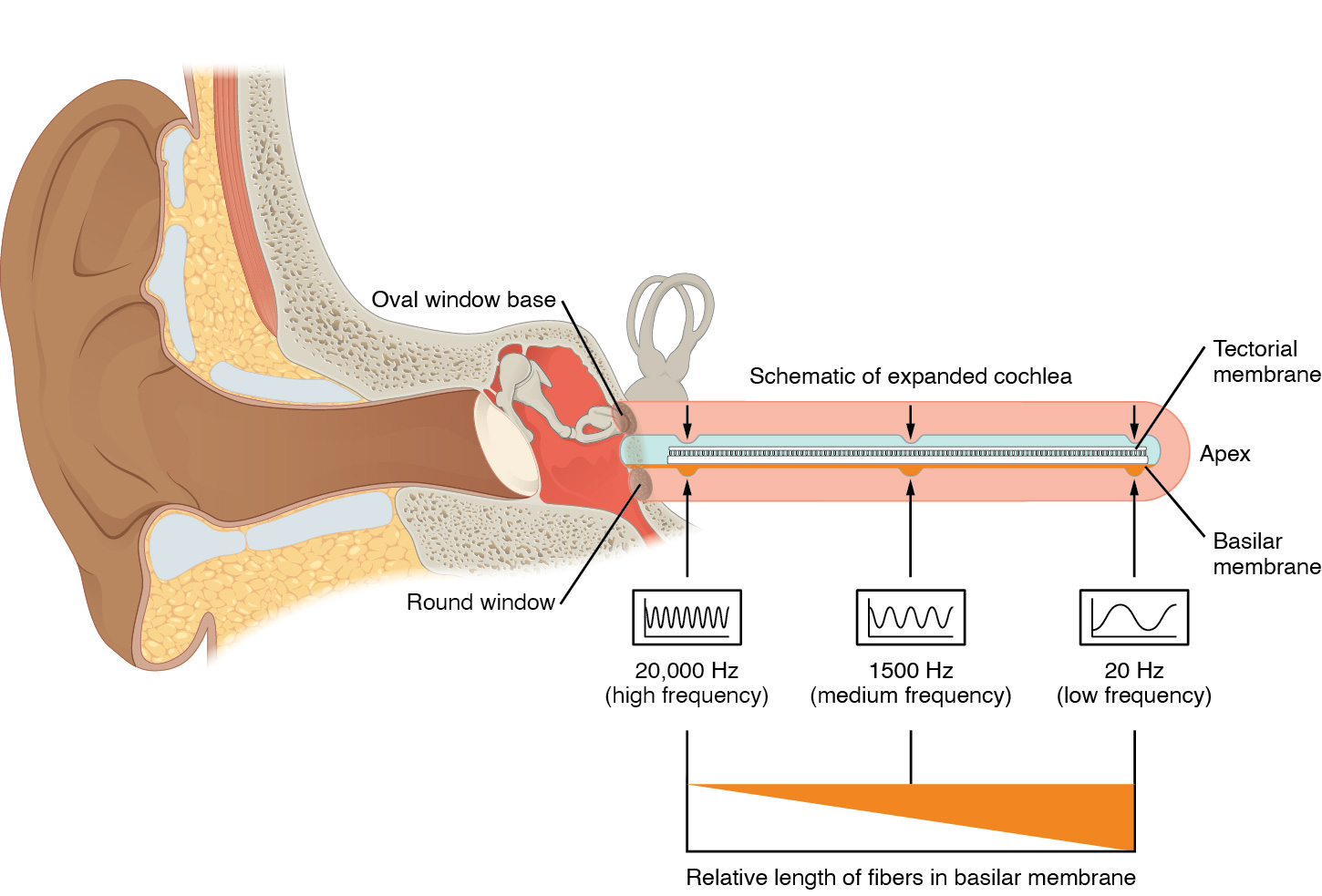
Kokleanın enine kesit görünümü, skala vestibuli ve skala timpani'nin koklear kanalın her iki tarafı boyunca uzandığını gösterir (aşağıdaki şekil). Koklear kanal, iki skalanın dalga hareketini nöral sinyallere dönüştüren birkaç Corti organı içerir. Corti organları, Corti organları ile skala timpani arasında bulunan koklear kanalın tarafı olan baziler zarın üstünde yer alır. Sıvı dalgaları skala vestibuli ve skala timpani boyunca hareket ederken, baziler zar dalgaların frekansına bağlı olarak belirli bir noktada hareket eder. Daha yüksek frekanslı dalgalar baziler zarın koklea tabanına yakın olan bölgesini hareket ettirir. Düşük frekanslı dalgalar baziler zarın kokleanın ucuna yakın olan bölgesini hareket ettirir.

Corti organları, hücrenin apikal yüzeylerinden uzanan saç benzeri stereosilyalar nedeniyle adlandırılan saç hücreleri içerir (aşağıdaki şekil). Stereosilya, uzundan kısaya doğru sıralanmış mikrovillus benzeri yapılardan oluşan bir dizidir. Protein lifleri her bir dizi içinde bitişik kılları birbirine bağlar, böylece dizi baziler zarın hareketlerine yanıt olarak bükülür. Stereosilya, tüy hücrelerinden Corti organının medialine bağlı olan üstteki tektoriyal zara kadar uzanır. Skaladan gelen basınç dalgaları baziler zarı hareket ettirdiğinde, tektoriyal zar stereosilya boyunca kayar. Bu, stereosilyaları her bir dizinin en uzun üyesine doğru ya da ondan uzağa doğru büker. Stereosilyalar dizilerinin en uzun üyesine doğru büküldüğünde, protein bağlarındaki gerilim saç hücresi zarındaki iyon kanallarını açar. Bu, tüy hücresi zarını depolarize edecek ve tüy hücrelerine bağlı afferent sinir lifleri boyunca ilerleyen sinir uyarılarını tetikleyecektir. Stereosilyalar dizilerinin en kısa üyesine doğru büküldüğünde, bağlar üzerindeki gerilim gevşer ve iyon kanalları kapanır. Ses olmadığında ve stereosilya düz durduğunda, bağlar üzerinde hala az miktarda gerilim vardır ve bu da tüy hücresinin zar potansiyelini hafifçe depolarize tutar.


| İNTERAKTİF BAĞLANTI Doku örneğini daha ayrıntılı incelemek için Michigan Üniversitesi WebScope'u görüntüleyin. Baziler zar, kokleanın merkezi çekirdeğinden kenarına doğru uzanan ince zardır. Koklea içindeki sıvıların hareketiyle aktive olabilmeleri için bu zara ne bağlanır? |
Yukarıda belirtildiği gibi, baziler zarın belirli bir bölgesi yalnızca gelen ses belirli bir frekanstaysa hareket edecektir. Tektoriyal zar sadece baziler zarın hareket ettiği yerde hareket ettiğinden, bu bölgedeki tüy hücreleri de sadece bu spesifik frekanstaki seslere yanıt verecektir. Bu nedenle, bir sesin frekansı değiştikçe, baziler zar boyunca farklı kıl hücreleri aktive olur. Koklea, insan kulağının algılayabildiği ses aralığı olan 20 ila 20.000 Hz arasındaki frekanslar için işitsel uyaranları kodlar. Hertz birimi, ses dalgalarının frekansını saniyede üretilen döngü cinsinden ölçer. 20Hz kadar düşük frekanslar, kokleanın tepesindeki veya ucundaki tüy hücreleri tarafından algılanır. Daha yüksek 20 KHz aralığındaki frekanslar, kokleanın tabanında, yuvarlak ve oval pencerelere yakın tüy hücreleri tarafından kodlanır (aşağıdaki şekil). Çoğu işitsel uyaran, çeşitli frekanslarda ve yoğunluklarda (ses dalgasının genliği ile temsil edilir) bir ses karışımı içerir. Koklear kanalın uzunluğu boyunca yer alan ve her biri belirli bir frekansa duyarlı olan tüy hücreleri, tıpkı bir prizmanın görünür ışığı bileşen renklerine ayırması gibi kokleanın da işitsel uyaranları frekansa göre ayırmasını sağlar.
| İNTERAKTİF BAĞLANTI Kulak yapılarının, koklear kanalın "kıllarını" veya stereosilyalarını hareket ettirerek ses dalgalarını nasıl sinirsel bir sinyale dönüştürdüğü hakkında daha fazla bilgi edinmek için bu videoyu izleyin. Kanalın uzunluğu boyunca belirli konumlar belirli frekansları veya perdeleri kodlar. Beyin, duyduğumuz seslerin anlamını müzik, konuşma, gürültü vb. olarak yorumlar. Sesin dış kulaktan iç kulağa amplifikasyonu ve transferinden hangi kulak yapıları sorumludur? |
| İNTERAKTİF BAĞLANTI İç kulak hakkında daha fazla bilgi edinmek ve tabanı görüntünün arkasında, tepesi ise önünde olacak şekilde kokleanın açıldığını görmek için bu animasyonu izleyin. Sesin belirli dalga boyları, tıpkı bir piyanonun tuşlarının farklı frekanslarda ses üretmesi gibi, baziler zarın belirli bölgelerinin titreşmesine neden olur. Animasyona dayanarak, yüksek perdeden alçak perdeye kadar frekanslar koklear kanal içindeki tüy hücrelerinde nerede aktiviteye neden olur? |
Denge (Balans)
İşitme duyusunun yanı sıra, iç kulak denge duyusu olan denge ile ilgili bilgilerin kodlanmasından sorumludur. Benzer bir mekanoreseptör -stereosilya içeren bir saç hücresi- baş pozisyonunu, baş hareketini ve vücudumuzun hareket halinde olup olmadığını algılar. Bu hücreler iç kulağın vestibülünde yer alır. Başın pozisyonu utrikül ve sakkül tarafından algılanırken, başın hareketi yarım daire kanalları tarafından algılanır. Vestibüler ganglionda üretilen nöral sinyaller vestibülokoklear sinir yoluyla beyin sapına ve beyinciğe iletilir.
Utrikül ve sakkülün her ikisi de büyük ölçüde makula dokusundan oluşur. Makula, destek hücreleri tarafından çevrelenmiş tüy hücrelerinden oluşur. Tüy hücrelerinin stereosilyaları otolitik zar adı verilen viskoz bir jelin içine uzanır (aşağıdaki şekil). Otolitik zarın üzerinde otolit adı verilen kalsiyum karbonat kristallerinden oluşan bir tabaka bulunur. Otolitler esasen otolitik zarı üst-ağır yapar. Otolitik zar, baş hareketlerine yanıt olarak makuladan ayrı hareket eder. Başın eğilmesi otolitik zarın yerçekimi yönünde makula üzerinde kaymasına neden olur. Hareket eden otolitik zar da sterosilyaları bükerek bazı tüy hücrelerinin depolarize olurken diğerlerinin hiperpolarize olmasına neden olur. Başın tam konumu beyin tarafından saç hücresi depolarizasyon modeline göre yorumlanır.

Yarım daire kanalları, vestibülün halka benzeri üç uzantısıdır. Biri yatay düzlemde, diğer ikisi ise dikey düzlemde yer alır. Ön ve arka dikey kanallar sagittal düzleme göre yaklaşık 45 derecelik bir açıyla yönlendirilmiştir (aşağıdaki şekil). Her bir yarım daire kanalının tabanı, vestibül ile birleştiği yerde, ampulla olarak bilinen genişlemiş bir bölgeye bağlanır. Ampulla, "hayır" derken başı çevirmek gibi dönme hareketine yanıt veren tüy hücrelerini içerir. Bu tüy hücrelerinin stereosilyaları, ampullanın tepesine yapışan bir zar olan kupulaya uzanır. Baş yarım daire kanalına paralel bir düzlemde dönerken, sıvı gecikir ve kupulayı baş hareketinin tersi yönde saptırır. Yarım daire kanalları birkaç ampulla içerir, bazıları yatay, diğerleri ise dikey yönelimlidir. Vestibüler sistem, hem yatay hem de dikey ampullanın göreceli hareketlerini karşılaştırarak, üç boyutlu uzaydaki çoğu baş hareketinin yönünü tespit edebilir.

Somatosensasyon (Dokunma)
Somatosensasyon, bu bölümde tartışılan özel duyuların aksine genel bir duyu olarak kabul edilir. Somatosensasyon, dokunma, propriyosepsiyon ve interosepsiyon ile ilişkili duyusal modaliteler grubudur. Bu modaliteler basınç, titreşim, hafif dokunma, gıdıklama, kaşıntı, sıcaklık, ağrı, propriyosepsiyon ve kinesteziyi içerir. Bu, reseptörlerinin özel bir organla ilişkili olmadığı, bunun yerine çeşitli organlarda vücuda yayıldığı anlamına gelir. Somatosensör reseptörlerinin çoğu deride bulunur, ancak reseptörler kaslarda, tendonlarda, eklem kapsüllerinde, bağlarda ve viseral organların duvarlarında da bulunur.
Serbest sinir uçları tarafından iletilen iki tür somatosensoriyel sinyal ağrı ve sıcaklıktır. Bu iki modalite, sıcaklık ve ağrı uyaranlarını iletmek için sırasıyla termoreseptörleri ve nosiseptörleri kullanır. Sıcaklık reseptörleri, yerel sıcaklıklar vücut sıcaklığından farklı olduğunda uyarılır. Bazı termoreseptörler sadece soğuğa, bazıları ise sadece sıcağa duyarlıdır. Nosisepsiyon, potansiyel olarak zarar verici uyaranların hissedilmesidir. Belirli bir eşiğin ötesindeki mekanik, kimyasal veya termal uyaranlar ağrılı hisleri ortaya çıkaracaktır. Stresli veya hasarlı dokular, nosiseptörlerdeki reseptör proteinlerini aktive eden kimyasallar salgılar. Örneğin, baharatlı yiyeceklerle ilişkili acı hissi, acı biberlerdeki aktif molekül olan kapsaisin içerir. Kapsaisin molekülleri, nosiseptörlerde 37°C'nin üzerindeki sıcaklıklara duyarlı bir transmembran iyon kanalına bağlanır. Kapsaisinin bu transmembran iyon kanalına bağlanma dinamiği, molekülün uzun süre bağlı kalması açısından olağandışıdır. Bu nedenle, diğer uyaranların aktive olmuş nosiseptör aracılığıyla ağrı hissi yaratma kabiliyetini azaltacaktır. Bu nedenle kapsaisin, Icy Hot™ gibi ürünlerde olduğu gibi topikal bir analjezik olarak kullanılabilir.
Parmağınızı dokulu bir yüzey üzerinde sürüklerseniz, parmağınızın derisi titreşecektir. Bu tür düşük frekanslı titreşimler, tip I kutanöz mekanoreseptörler olarak da bilinen Merkel hücreleri adı verilen mekanoreseptörler tarafından algılanır. Merkel hücreleri epidermisin stratum bazalesinde bulunur. Derin basınç ve titreşim, dermisin veya deri altı dokusunun derinliklerinde bulunan kapsüllü uçlara sahip reseptörler olan lamelli (Pacinian) korpüsküller tarafından iletilir. Hafif dokunma, dokunsal (Meissner) korpüsküller olarak bilinen kapsüllenmiş uçlar tarafından iletilir. Foliküller ayrıca saç folikülü pleksusu olarak bilinen bir sinir uçları pleksusu ile sarılmıştır. Bu sinir uçları, bir böceğin deri üzerinde yürümesi gibi, deri yüzeyindeki tüylerin hareketini algılar. Derinin gerilmesi, soğansı cisimcikler olarak bilinen gerilme reseptörleri tarafından iletilir. Bulböz korpüsküller, Ruffini korpüskülleri veya tip II kutanöz mekanoreseptörler olarak da bilinir.
Diğer somatosensoriyel reseptörler eklemlerde ve kaslarda bulunur. Esneme reseptörleri tendonların, kasların ve eklem bileşenlerinin gerilmesini izler. Örneğin, hiç egzersizden önce veya sonra kaslarınızı gerdiniz ve kaslarınız daha az gerilmiş bir duruma geri spazm geçirmeden önce sadece bir yere kadar esneyebildiğinizi fark ettiniz mi? Bu spazm, kas yırtılmasını önlemek için gerilme reseptörleri tarafından başlatılan bir reflekstir. Bu tür gerilme reseptörleri bir kasın aşırı kasılmasını da önleyebilir. İskelet kası dokusunda bu gerilme reseptörlerine kas iğcikleri adı verilir. Golgi tendon organları da benzer şekilde tendonların gerilme seviyelerini aktarır. Bulböz korpüsküller eklem kapsüllerinde de bulunur ve burada eklem içindeki iskelet sistemi bileşenlerindeki gerilmeyi ölçerler. Sinir uçlarının türleri, konumları ve ilettikleri uyarılar aşağıdaki tabloda sunulmuştur.
| İsim | Tarihsel isim | Konum(lar) | Uyaranlar |
| Serbest sinir uçları | * | Dermis, kornea, dil, eklem kapsülleri, viseral organlar | Ağrı, sıcaklık, mekanik deformasyon |
| Mekanoreseptörler | Merkel’in diskleri | Epidermal-dermal bileşke, mukozal zarlar | Düşük frekanslı titreşim (5-15 Hz) |
| Bulbous korpüskül | Ruffini’nin korpüskülü | Dermis, eklem kapsülleri | Esneme |
| Dokunsal korpüskül | Meissner’in korpüskülü | Papiller dermis, özellikle parmak uçlarında ve dudaklarda | Hafif dokunuş, 50 Hz’in altında titreşimler |
| Lamelli korpüskül | Pacinian korpüskülü | Derin dermis, deri altı doku | Derin basınç, yüksek frekanslı titreşim (yaklaşık 250 Hz) |
| Saç folikülü pleksusu | * | Dermisteki saç foliküllerinin etrafına sarılır | Kıl hareketleri |
| Kas mili | * | İskelet kası lifleri ile uyumlu | Kas kasılması ve gerilmesi |
| Tendon germe organı | Golgi tendon organı | Tendonlar ile aynı hizada | Tendonların gerilmesi |
Vizyon
Görme, gözler aracılığıyla alınan ışık uyaranlarının transdüksiyonuna dayanan özel bir duyudur. Gözler kafatasında her iki yörüngenin içinde yer alır. Kemik orbita göz kürelerini çevreleyerek onları korur ve gözün yumuşak dokularını sabitler (aşağıdaki şekil). Ön kenarlarında kirpikler bulunan göz kapakları, göz yüzeyine gelebilecek parçacıkları engelleyerek gözün aşınmalara karşı korunmasına yardımcı olur. Her bir kapağın iç yüzeyi palpebral konjonktiva olarak bilinen ince bir zardır. Konjonktiva gözün beyaz bölgeleri (sklera) üzerinde uzanır ve göz kapaklarını göz küresine bağlar. Gözyaşı, orbitanın hemen içinde, göz küresinin üst ve yan tarafında bulunan lakrimal bez tarafından üretilir. Bu bez tarafından üretilen gözyaşları lakrimal kanaldan gözün medial köşesine akar ve burada gözyaşları konjonktiva üzerinden akarak yabancı partikülleri yıkar.

Gözün orbita içindeki hareketi, orbita kemiklerinden çıkan ve göz küresinin yüzeyine yerleşen altı ekstraoküler kasın kasılmasıyla gerçekleşir (aşağıdaki şekil). Kasların dördü gözün etrafındaki ana noktalarda düzenlenmiştir ve bu konumlara göre adlandırılmıştır. Bunlar superior rektus, medial rektus, inferior rektus ve lateral rektustur. Bu kasların her biri kasıldığında, göz kasılmakta olan kasa doğru hareket eder. Örneğin, üst rektus kasıldığında, göz yukarı bakmak için döner. Superior oblik, arka yörüngede, dört rektus kasının orijinine yakın bir yerde ortaya çıkar. Bununla birlikte, eğik kasların tendonu troklea olarak bilinen kasnak benzeri bir kıkırdak parçasından geçer. Tendon, gözün üst yüzeyine eğik bir şekilde yerleşir. Tendonun troklea ile yaptığı açı, superior oblik kasının kasılmasının gözü lateral olarak döndürdüğü anlamına gelir. İnferior oblik kas orbita tabanından kaynaklanır ve gözün inferolateral yüzeyine eklenir. Kasıldığında, süperior oblik kasın tersine gözü lateral olarak döndürür. Gözün iki oblik kas tarafından döndürülmesi gereklidir çünkü göz sagittal düzlemde mükemmel bir şekilde hizalanmamıştır. Göz yukarı ya da aşağı baktığında, üst rektusun düz yukarı yerine yaklaşık 20 derecelik bir açıyla çekmesini telafi etmek için gözün de hafifçe dönmesi gerekir. Aynı durum inferior oblik kasın kasılmasıyla telafi edilen inferior rektus için de geçerlidir. Orbitadaki yedinci kas, üst göz kapağının kaldırılıp geri çekilmesinden sorumlu olan levator palpebrae superioris'tir ve bu hareket genellikle gözün superior rektus tarafından kaldırılmasıyla birlikte gerçekleşir (yukarıdaki şekil).
Ekstraoküler kaslar üç kraniyal sinir tarafından innerve edilir. Gözün abdüksiyonuna neden olan lateral rektus, abdusens siniri tarafından innerve edilir. Superior oblik troklear sinir tarafından innerve edilir. Levator palpebrae superioris gibi diğer tüm kaslar okülomotor sinir tarafından innerve edilir. Bu kraniyal sinirlerin motor çekirdekleri, göz hareketlerini koordine eden beyin sapına bağlanır.

Gözün kendisi üç doku katmanından oluşan içi boş bir küredir. En dıştaki katman, beyaz sklera ve şeffaf korneayı içeren fibröz tuniktir. Sklera, göz yüzeyinin altıda beşini oluşturur ve çoğu görünmez, ancak insanlar "gözün beyazının" bu kadar büyük bir kısmının görünür olmasıyla diğer birçok türe kıyasla benzersizdir (aşağıdaki şekil). Şeffaf kornea gözün ön ucunu kaplar ve ışığın göze girmesini sağlar. Gözün orta tabakası, çoğunlukla koroid, siliyer cisim ve iristen oluşan vasküler tuniktir. Koroid, göz küresine kan akışını sağlayan yüksek oranda vaskülarize olmuş bir bağ dokusu tabakasıdır. Koroid, askı bağları veya zonül lifleri ile lense bağlanan kaslı bir yapı olan siliyer cismin arkasındadır. Bu iki yapı lensi bükerek ışığın gözün arka tarafına odaklanmasını sağlar. Siliyer cismin üzerinde yer alan ve gözün ön kısmında görülebilen iris, gözün renkli kısmıdır. İris, gözün merkezinde ışığın girmesini sağlayan delik olan göz bebeğini açan veya kapatan düz bir kastır. İris, parlak ışığa tepki olarak göz bebeğini daraltır ve loş ışığa tepki olarak göz bebeğini genişletir. Gözün en iç tabakası, fotoresepsiyondan sorumlu sinir dokusunu içeren nöral tunik veya retinadır.
Göz ayrıca iki boşluğa ayrılır: ön boşluk ve arka boşluk. Ön boşluk, iris ve siliyer cisim de dahil olmak üzere kornea ve lens arasındaki boşluktur. Aköz hümör adı verilen sulu bir sıvı ile doludur. Arka boşluk, lensin arkasında retinanın bulunduğu iç göz küresinin arka tarafına uzanan boşluktur. Arka boşluk vitreus humor adı verilen daha viskoz bir sıvı ile doludur.
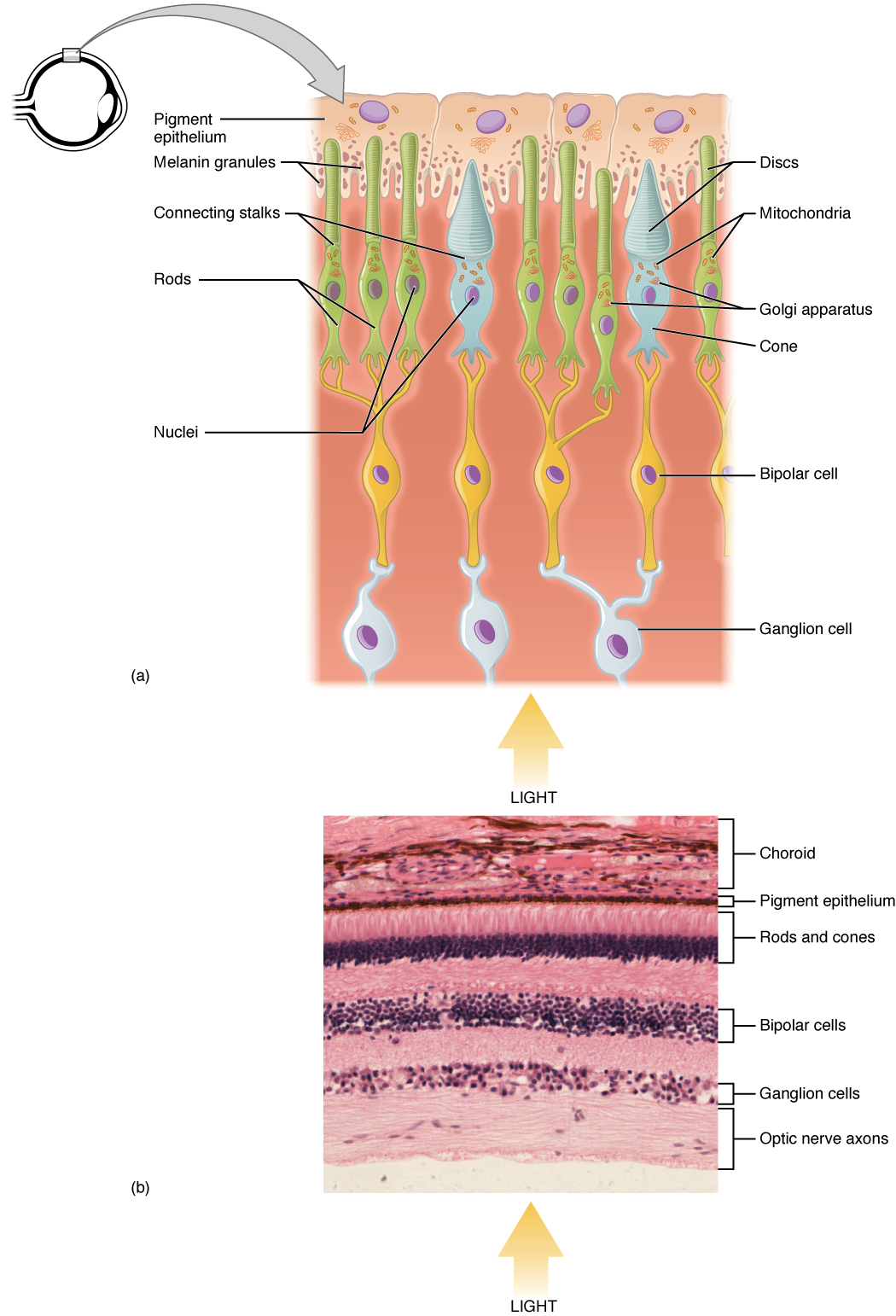
Retina birkaç katmandan oluşur ve görsel uyaranların ilk işlenmesi için özelleşmiş hücreler içerir. Fotoreseptörler (çubuklar ve koniler) ışık enerjisi ile uyarıldıklarında membran potansiyellerini değiştirirler. Zar potansiyelindeki değişiklik, fotoreseptör hücrelerinin dış sinaptik katmandaki bipolar hücrelere saldığı nörotransmitter miktarını değiştirir. Bir fotoreseptörü iç sinaptik katmandaki retinal ganglion hücresine (RGC) bağlayan retinadaki bipolar hücredir. Burada, amprin hücreleri ayrıca RGC tarafından bir aksiyon potansiyeli üretilmeden önce retinal işleme katkıda bulunur. Retinanın en iç tabakasında bulunan RGC'lerin aksonları optik diskte toplanır ve optik sinir olarak gözü terk eder (aşağıdaki şekil). Bu aksonlar retinadan geçtiği için, gözün en arkasında, optik sinirin başladığı yerde fotoreseptör yoktur. Bu durum retinada bir "kör nokta" ve görme alanımızda da buna karşılık gelen bir kör nokta oluşturur.

Retinadaki fotoreseptörlerin (çubuklar ve koniler) aksonların, RGC'lerin, bipolar hücrelerin ve retinal kan damarlarının arkasında yer aldığını unutmayın. Işığın önemli bir kısmı, ışık fotoreseptör hücrelerine ulaşmadan önce bu yapılar tarafından emilir. Bununla birlikte, retinanın tam merkezinde fovea olarak bilinen küçük bir alan bulunur. Foveada retina destek hücreleri ve kan damarlarından yoksundur ve sadece fotoreseptörler içerir. Bu nedenle, görme keskinliği veya görme netliği foveada en yüksek seviyededir. Bunun nedeni foveanın gelen ışığın diğer retinal yapılar tarafından en az emildiği yer olmasıdır (yukarıdaki şekil). Retinanın bu merkezi noktasından herhangi bir yöne doğru hareket edildiğinde, görme keskinliği önemli ölçüde azalır. Ayrıca, foveanın her bir fotoreseptör hücresi tek bir RGC'ye bağlıdır. Bu nedenle, bu RGC'nin birden fazla fotoreseptörden gelen girdileri entegre etmesi gerekmez, bu da görsel iletimin doğruluğunu azaltır. Retinanın kenarlarına doğru, birkaç fotoreseptör RGC'lere (bipolar hücreler aracılığıyla) 50'ye 1 oranına kadar yaklaşır. Fovea ve periferik retina arasındaki görme keskinliği farkı, bu paragrafın ortasındaki bir kelimeye doğrudan bakılarak kolayca kanıtlanabilir. Görüş alanının ortasındaki görsel uyaran fovea üzerine düşer ve en keskin odaktadır. Gözlerinizi o kelimeden ayırmadan, paragrafın başındaki veya sonundaki kelimelerin odakta olmadığına dikkat edin. Çevrenizdeki görüntüler, periferik retina tarafından odaklanır ve belirsiz, bulanık kenarlara sahiptir ve kelimeler net bir şekilde tanımlanmaz. Bu nedenle, gözlerin büyük bir sinirsel fonksiyonu, önemli görsel uyarıların fovea üzerinde merkezlenmesi için gözleri ve başın hareket ettirilmesiyle ilgilidir.
Retina üzerine düşen ışık, fotoreseptörlerdeki pigment moleküllerinde kimyasal değişikliklere neden olur ve sonuçta RGC'lerin aktivitesinde bir değişikliğe yol açar. Fotoreseptör hücreleri iç segment ve dış segment olmak üzere iki bölümden oluşur (aşağıdaki şekil). İç segment bir hücrenin çekirdeğini ve diğer ortak organellerini içerirken, dış segment fotoresepsiyonun gerçekleştiği özel bir bölgedir. Dış segmentlerinin şekli bakımından farklılık gösteren iki tür fotoreseptör vardır -çubuklar ve koniler-. Çubuk fotoreseptörün çubuk şeklindeki dış segmentleri, ışığa duyarlı pigment rodopsin içeren membrana bağlı disklerden oluşan bir yığın içerir. Koni fotoreseptörünün koni şeklindeki dış segmentleri, ışığa duyarlı pigmentlerini hücre zarının katlanmalarında içerir. Opsin adı verilen ve her biri ışığın belirli bir dalga boyuna duyarlı olan üç koni fotopigmenti vardır. Görünür ışığın dalga boyu rengini belirler. İnsan gözündeki pigmentler üç farklı ana rengi algılamak üzere özelleşmiştir: kırmızı, yeşil ve mavi.
Moleküler düzeyde, görsel uyaranlar fotopigment molekülünde değişikliklere neden olur ve bu da fotoreseptör hücrenin zar potansiyelinde değişikliklere yol açar. Tek bir ışık birimine foton denir ve fizikte hem parçacık hem de dalga özelliklerine sahip bir enerji paketi olarak tanımlanır. Bir fotonun enerjisi dalga boyu ile temsil edilir ve görünür ışığın her dalga boyu belirli bir renge karşılık gelir. Görünür ışık, dalga boyu 380 ile 720 nm arasında olan elektromanyetik radyasyondur. Elektromanyetik radyasyonun 720 nm'den uzun dalga boyları kızılötesi aralığa girerken, 380 nm'den kısa dalga boyları ultraviyole aralığına girer. Dalga boyu 380 nm olan ışık mavi iken, dalga boyu 720 nm olan ışık koyu kırmızıdır. Diğer tüm renkler, dalga boyu skalası boyunca çeşitli noktalarda kırmızı ve mavi arasında yer alır.
Opsin pigmentleri aslında retinal olarak bilinen bir kofaktör içeren transmembran proteinlerdir. Retinal, A vitamini ile ilişkili bir hidrokarbon molekülüdür. Bir foton retinale çarptığında, molekülün uzun hidrokarbon zinciri biyokimyasal olarak değişime uğrar. Özellikle, fotonlar zincirdeki çift bağlı karbonlardan bazılarının cis'ten trans konformasyonuna geçmesine neden olur. Bu sürece fotoizomerizasyon denir. Bir fotonla etkileşime girmeden önce, retinalin esnek çift bağlı karbonları cis konformasyonundadır. Bu molekül 11-cis-retinal olarak adlandırılır. Molekülle etkileşime giren bir foton, esnek çift bağlı karbonların trans- konformasyona geçmesine neden olarak düz bir hidrokarbon zincirine sahip all-trans-retinal oluşturur (aşağıdaki şekil).
Fotoreseptörlerdeki retinalin şekil değiştirmesi retinada görsel iletimi başlatır. Retinal ve opsin proteinlerinin aktivasyonu bir G proteininin aktivasyonuyla sonuçlanır. G proteini fotoreseptör hücresinin zar potansiyelini değiştirerek retinanın dış sinaptik tabakasına daha az nörotransmitter salgılar. Retina molekülü 11-cis-retinal şekline geri dönene kadar, opsin ışık enerjisine yanıt veremez, buna ağartma denir. Büyük bir fotopigment grubu ağartıldığında, retina sanki karşıt görsel bilgi algılanıyormuş gibi bilgi gönderecektir. Parlak bir ışık parlamasından sonra, art görüntüler genellikle negatif olarak görülür. Fotoizomerizasyon bir dizi enzimatik değişiklikle tersine çevrilir, böylece retinal daha fazla ışık enerjisine yanıt verir.

Opsinler ışığın sınırlı dalga boylarına duyarlıdır. Rodoplardaki fotopigment olan rodopsin, en çok 498 nm dalga boyundaki ışığa duyarlıdır. Üç renk opsini 564 nm, 534 nm ve 420 nm'lik pik hassasiyetlere sahiptir ve bu da kabaca kırmızı, yeşil ve mavinin ana renklerine karşılık gelir (aşağıdaki şekil). Rodopsinin çubuklardaki absorbansı koni opsinlerinden çok daha hassastır; özellikle çubuklar düşük ışık koşullarında görmeye duyarlıdır ve koniler daha parlak koşullara duyarlıdır. Normal güneş ışığında, koniler aktifken rodopsin sürekli olarak ağaracaktır. Karanlık bir odada, koni opsinlerini aktive etmek için yeterli ışık yoktur ve görme tamamen çubuklara bağlıdır. Çubuklar ışığa o kadar duyarlıdır ki, tek bir foton bir çubuğun ilgili RGC'sinde bir aksiyon potansiyeline yol açabilir.
Işığın farklı dalga boylarına duyarlı olan üç tip koni opsini, renkli görmemizi sağlar. Beyin, üç farklı koninin aktivitesini karşılaştırarak görsel uyaranlardan renk bilgisi çıkarabilir. Örneğin, yaklaşık 450 nm dalga boyuna sahip parlak mavi bir ışık "kırmızı" konileri minimum düzeyde, "yeşil" konileri marjinal düzeyde ve "mavi" konileri ağırlıklı olarak aktive edecektir. Üç farklı koninin göreceli aktivasyonu, rengi mavi olarak algılayan beyin tarafından hesaplanır. Bununla birlikte, koniler düşük yoğunluklu ışığa tepki veremez ve çubuklar ışığın rengini algılamaz. Bu nedenle, düşük ışıkta görüşümüz özünde gri tonlamalıdır. Başka bir deyişle, karanlık bir odada her şey grinin bir tonu olarak görünür. Karanlıkta renkleri görebildiğinizi düşünüyorsanız, bunun nedeni büyük olasılıkla beyninizin bir şeyin ne renk olduğunu bilmesi ve bu hafızaya güvenmesidir.
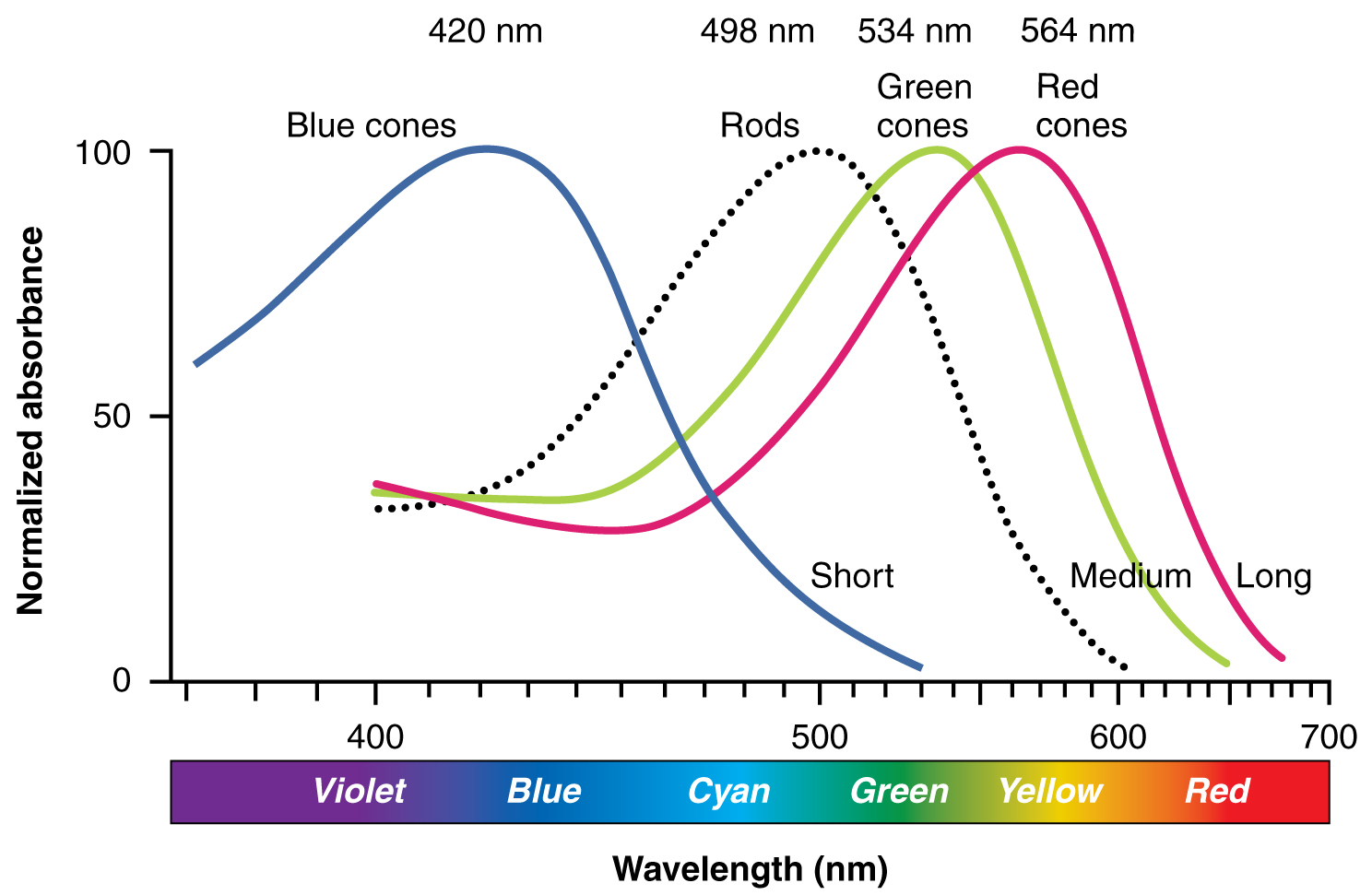
Fotopigmentlerin Renk Hassasiyetinin Karşılaştırılması Dört fotopigmentin pik hassasiyeti ve absorbans spektrumlarının karşılaştırılması, bunların belirli dalga boylarına karşı en hassas olduklarını göstermektedir.
| İNTERAKTİF BAĞLANTI Gözden oksipital kortekse giden görsel yolu gösteren beynin enine kesiti hakkında daha fazla bilgi edinmek için bu videoyu izleyin. Yolun ilk yarısı, RGC'lerden optik sinir yoluyla her iki taraftaki talamusta bulunan lateral genikülat çekirdeğe olan projeksiyondur. Bu yoldaki ilk lif, talamik bir hücre üzerinde sinaps yapar ve daha sonra "görmenin" veya görsel algının gerçekleştiği oksipital lobdaki görsel kortekse projekte olur. Bu video, gözlerden oksipital loba giden yola odaklanarak görme sistemine kısaltılmış bir genel bakış sunmaktadır. Videoda (0:45'te) "retinadaki ganglion hücreleri adı verilen özelleşmiş hücreler ışık ışınlarını elektrik sinyallerine dönüştürür" ifadesi yer almaktadır. Bu ifade retinal işlemenin hangi yönünü basitleştirmektedir? Cevabınızı açıklayınız. |
Duyusal Sinirler
Herhangi bir duyu hücresi bir uyaranı sinir impulsuna dönüştürdüğünde, bu impulsun MSS'ye ulaşmak için aksonlar boyunca ilerlemesi gerekir. Özel duyuların çoğunda, duyusal reseptörleri terk eden aksonlar topografik bir düzenlemeye sahiptir, yani duyusal reseptörün konumu aksonun sinirdeki konumuyla ilgilidir. Örneğin, retinada, foveadaki RGC'lerden gelen aksonlar optik sinirin merkezinde bulunur ve burada daha periferik RGC'lerden gelen aksonlarla çevrilidir.
Omurilik Sinirleri
Genel olarak spinal sinirler, deri gibi periferdeki duyusal reseptörlerden gelen afferent aksonlar ile kaslara veya diğer efektör organlara giden efferent aksonları içerir. Spinal sinir omuriliğe yaklaştıkça dorsal ve ventral köklere ayrılır. Dorsal kök sadece duyusal nöronların aksonlarını içerirken, ventral kökler sadece motor nöronların aksonlarını içerir. Bazı dallar, girdikleri omurilik seviyesinde dorsal kök gangliyonundaki, arka (dorsal) boynuzdaki ve hatta ön (ventral) boynuzdaki yerel nöronlarla sinaps yapacaktır. Diğer dallar omuriliğin diğer seviyelerindeki nöronlarla etkileşime girmek için omurganın yukarısına veya aşağısına doğru kısa bir mesafe kat eder. Bir dal beyne bağlanmak için beyaz maddenin arka (dorsal) kolonuna da dönebilir. Kolaylık olması açısından, bu yolların bir parçası olan omurilik içindeki yapılara atıfta bulunmak için ventral ve dorsal terimlerini kullanacağız. Bu, farklı bileşenler arasındaki ilişkilerin altını çizmeye yardımcı olacaktır. Tipik olarak, beyne bağlanan spinal sinir sistemleri kontralateraldir, yani vücudun sağ tarafı beynin sol tarafına ve vücudun sol tarafı beynin sağ tarafına bağlıdır.
Kraniyal Sinirler
Kraniyal sinirler baş ve boyundan gelen belirli duyusal bilgileri doğrudan beyne iletir. Spinal bilgi kontralateral iken, bazı istisnalar dışında kraniyal sinir sistemleri çoğunlukla ipsilateraldir, yani başın sağ tarafındaki bir kraniyal sinir beynin sağ tarafına bağlıdır. Koku alma, optik ve vestibülokoklear sinirler gibi bazı kraniyal sinirler yalnızca duyusal aksonlar içerir. Trigeminal, fasiyal, glossofaringeal ve vagus sinirleri de dahil olmak üzere diğer kraniyal sinirler hem duyusal hem de motor aksonlar içerir (ancak vagus siniri somatik sinir sistemi ile ilişkili değildir). Yüz için genel somatosensasyon duyuları trigeminal sistemden geçer.
Önceki Ders: Somatik Sinir Sistemi (Başlangıç)

Yorumlar
Yorum Gönder